ADDRESSING
VOCALREGISTER DISCREPANCIES:
AN ALTERNATIVE,
SCIENCE-BASED THEORY
OF REGISTER PHENOMENA
Leon
Thurman, Ed.D.
Graham
Welch, Ph.D.
Axel
Theimer, D.M.A.
Carol Klitzke, M.S., CCC/SLP
Second International Conference
The Physiology and Acoustics of Singing
6 – 9 October 2004
Abstract
This paper analyzes and evaluates several
explicit and implicit assumptions that are embedded in the concepts,
terminologies, and practices related to vocal registers. A reconciliation of the varied and
conflicting register concepts, terminologies, and practices is proposed,
including:
1.
a
brief historical context of vocal registers,
2.
a
documented science-based theory that accounts for vocal register phenomena from
perceptual, physiological, and acoustical perspectives,
3.
criteria
for selection of science-based categorical word labels for register phenomena
and suggested colloquial English terms that meet the criteria,
4.
how
the theory can be beneficially applied to the learning of efficient, skilled
singing and speaking when guided by music educators, choral conductors, singing
teachers, speech teachers, theatre directors, and when applied to therapeutic
clinical settings.
“You can't make me sing
that song. Those notes are past my break
and I'd have to use that weak part of my voice that just sounds awful. I won't do it.”
“I've always been a
soprano, so my lower pitch range is pretty weak. In fact, I was taught that singing in chest
register can damage your voice. I
certainly don't want to do that.”
“I've always been an alto
and when I sing pitches in the middle of the treble staff, my voice has a kind
of edgy, hard sound that sticks out in the choir. Then, above that, my voice is breathy and
weak. I've always wanted to be able to
sing with a high, clear soprano voice, but I can't.”
Male: “I can sing fine up
to about a D, and from G on up is sort of OK.
But right in between, my voice is real inconsistent. Sometimes it just gets weak, or it goes out of
tune, and sometimes it just flips or cracks.
Can you help me?”
“That Mariah Carey can
really sing high notes--and I mean high notes!
It's incredible. She was blessed
with a one-in-a-billion gift.”
Female: “I had one singing
teacher who called my head voice 'falsetto'.
Does that mean I'm supposed to sound like a man when he sings in his
falsetto?”
“What is my chest voice and
my head voice? And somebody told me
there is supposed to be a middle voice in between them. I'm confused.
Can you explain about my voices and show me when I'm supposed to be in
one or the other?”
“My choir director says
there are two voices--chest voice and head voice. My singing teacher talks about chest, middle,
and falsetto registers. I read a book on
voice that said there was a heavy mechanism and a light mechanism, whatever
those are. I wish you guys would make up
your minds; this is frustrating.”
Vocal registers
are controversial in the pedagogical, clinical, and scientific domains of
vocology. A well known general
definition of vocal registers is “...perceptually distinct regions of vocal quality that can be maintained over some ranges of
pitch and loudness.” (Titze, 2000, p. 282)
For centuries, however, concepts and practices related to vocal register
phenomena, including their linguistic labels, have been somewhat varied and
commonly contradictory.
Within both the
voice science and the voice education communities of the early 21st
century, considerable discrepancies remain in the conceptual frameworks,
terminologies, and practices that are related to vocal registers. To people who are not familiar with the
jargon of the voice-related professions, these discrepancies are puzzling,
confusing, and can even call into question the credibility of voice profession members.
In this paper, a
reconciliation of the varied and conflicting register concepts, terminologies,
and practices will be presented. It
will: (1) present a brief historical context of vocal registers,
(2) propose a
documented science-based theory that accounts for all vocal register phenomena
from perceptual, physiological, and acoustical perspectives, (3) propose
criteria for selection of categorical word labels for register phenomena and
suggest terms that meet them, and (4) suggest how the theory can be
beneficially applied to the learning of efficient, skilled singing and speaking
when guided by music educators, choral conductors, singing teachers, speech
teachers, theatre directors, and when applied to therapeutic clinical settings.
A BRIEF HISTORY OF VOCAL
REGISTERS
Perhaps for
millennia of time, singers and singing teachers have aurally identified changes
in voice quality when singing the consecutive fundamental frequencies (F0s) of a two-octave (or more) musical
scale. When transitions from one voice
quality to another occur, most singers report some sort of non-specific,
kinesthetically sensed, neuromuscular coordination adjustment in the
larynx. Among experienced or trained
singers, the transitions are perceived to be blended and smooth, but the
transitions are more commonly abrupt among inexperienced
singers.
The writings of
the Greek physician Galen (c. 129-200 AD) were the “bible” of medical anatomy
and practice for almost 1500 years after his death, but his observations
contained quite a number of inaccuracies.
Detailed knowledge of human anatomy and physiology began to be assembled
in the mid-16th century. A
prominent center for such study was the
Scientific
findings about vocal anatomy,
physiology, and the nature of vocal sound only began to be widely distributed
in about the middle of the 19th century.
With limited science-based knowledge of vocal anatomy, physiology, and
acoustics, singers and singing teachers had
to base their vocal concepts, terminologies, and practices substantially on
logical assumptions, personal perceptions, and metaphoric communications about
the nature of voices.
The term register was borrowed from the
terminology of keyboard organs (Merkel, 1863), and has been used in vocal
terminology since at least the 13th century (Duey, 1951). The earliest known writings about voice date
from that century. They were written in
Latin by two monks, Jerome of Moravia (c.1250) and John of Garland (c.1193 -
c.1270) (Large, 1973, p. 10; Mori, 1970; Timberlake, 1990). They wrote about the then current conceptual
categories and linguistic labels for the various “voices” in which singers can
sing.
When singers
sang in their upper pitch range, for instance, they presumably felt a
prominence of vibration sensations in the front, sides, and/or top of their
heads. We may presume that they
interpreted those sensations as meaning that their voices were “coming from”
their heads, so logically, they would call that way of singing head voice (Latin: vox captis = voice from the head).
When they perceived their head voice as producing subtle voice quality
differences, they are likely to have experienced differences in the vibration
sensations in their heads and believed they could “place” their head voices in
different areas of the head.
When they sang
in their lower pitch range, they are likely to have felt a prominence of
vibrations in their chest. We may
presume, therefore, that they interpreted those sensations as meaning that
their voices were “coming from” their chests and would call that way of singing
their chest voice (Latin: vox pectoris = voice from the breast or
chest). When they sang in their middle
pitch range, they no longer felt prominent vibration sensations in the head or
chest, but presumably felt a prominence of such sensations in their throats
because they called that way of singing their throat voice (Latin: vox
gutturis = voice in the throat).
When they changed from one voice to another, they physically felt the
transition from one “place” in the body to another, and they heard the sound quality of their voices change at the
same time. We may presume that this is
how a language of “voice placement” evolved.
In the 17th
century, Caccini wrote of voce piena
(full voice) and voce finta (feigned
voice). In 1627, Monteverdi wrote
of la
vocale della gola (the voice of the throat) and la vocale
In the 19th
century, Garcia and others wrote of three registers in ascending order of pitch
range: chest, falsetto, and head
(Garcia, 1855; also reported in Large, 1973, p.10; Timberlake, 1990). Merkel (1873), wrote of chest and falsetto
registers in men and low, middle, and high for women. In 1875,
John Curwen suggested thick, thin, and small as names for vocal registers (cited in Mackworth-Young,
1953). Browne and Behnke (1884) wrote
about five registers, i.e., lower thick,
upper thick, lower thin, upper thin,
and small. In the early 20th century,
Fröschels (1920) maintained that a “natural” voice had no registers, so that
voices were only one register. Wilcox
(1935) suggested the terms heavy
mechanism and light mechanism for
two vocal registers.
When singing in
their middle pitch range, some singers noticed sensations and sound qualities
that were different from those that they observed when singing in their
uppermost and lower pitch ranges. This
way of singing seemed to be a mixture of the sounds and sensations of chest and head or falsetto, so it
has come to be called a medium, or middle, or mixed register (Italian: voce
mista; French: voix mixte). When singing in middle or mixed voice,
singers observed transitions to other registers at both the top and bottom of
the mixed voice (Mori, 1970; Timberlake, 1990).
Its pitch range was said to be between the chest and the head or falsetto voices. According to Miller's description of the
Italian vocal pedagogy tradition (1977; 1986, pp. 115-149), the primo passaggio was the passage from
chest register to middle register and the secondo
passaggio was the passage from middle to head register. Men and women of various voice
classifications experienced the passages at slightly different pitch ranges.
Currently, the
terms head register and falsetto register are used by various
vocal pedagogues and voice scientists as labels for the different sound
qualities produced in middle and higher pitch ranges. For some singing teachers, however, head register is immediately above chest, and falsetto is above head. To
others, falsetto refers to all sound
qualities above chest in both males
and females, and sometimes the traditional high-range male falsetto is termed pure
falsetto. Among speakers of English,
the common, colloquial use of the
term falsetto refers only to the
female-like voice quality that males can make.
In some register
concepts, there are two “auxiliary registers”, one above and one below the more
commonly used head and chest registers.
Some prepubescent children, changing-voice males, and changed-voice male
and female adults can have a register that is variously labeled whistle, flute, or flageolet register that enables pitches that are
quite high (Cooksey, 2000, Large, 1973; Miller, 1986, pp. 147-148; Mori, 1970;
Timberlake, 1990). Some changed-voice
males and females are able to produce unusually low pitches below the more
commonly used registers. It has been
referred to as pulse or Strohbass register.
In the late
1960s, the research team of Minoru Hirano, William Vennard, and John Ohala (1969, 1970; Vennard, et al., 1970a,b)
heightened interest in the use of the scientific method to resolve many
controversies in the vocal pedagogy tradition, including landmark studies on
fundamental frequency, intensity, and register phenomena. They suggested Wilcox’s label heavy mechanism as a substitute for
chest register and his term light
mechanism as a substitute for registers above heavy
mechanism. In 1967, both Ralph Appelman
and William Vennard published landmark, science-based vocal pedagogy
books. In 1971, the late Wilbur James
Gould, M.D., founded the Voice Foundation in
In 1974, Harry
Hollien, an internationally prominent speech-voice scientist, presented four
new register terminologies for use in the speech science community, based on
his wide research experience:
1. Pulse register—a pulsated quality that
can be produced in a very low pitch range below modal, a sound quality that
also is called vocal fry.
2. Modal register—a heavier or thicker
voice quality that is produced in a lower pitch range. The label was a reference to the most common
"mode" of voice function, i.e., speech. It was the speech equivalent of chest
register in singing pedagogy.
3. Loft register—a voice quality that is
higher in pitch and lighter or thinner in quality, compared to modal register. It was the speech equivalent of head or
falsetto register in singing pedagogy.
4. Flute register—an even thinner quality
that can be produced in a very high pitch range above loft. In singing it is called falsetto in males and in females it is sometimes referred to as whistle register.
With increased
sophistication of instruments that are capable of documenting various vocal
phenomena came a curiosity about the actual anatomic, physiologic, and acoustic
realities of vocal registers and their transitions in both speaking and
singing. In the late 1970s, an
international medical organization, the Collegium Medicorum Theatri (CoMeT),
formed an international Committee on Vocal Registers with Dr. Hollien as
Chair. The committee included prominent
otolaryngologists, speech and voice scientists, and singing teachers. Their charge was to attempt a definition of
vocal registers perceptually, physiologically, and acoustically.
After their
early meetings, they agreed that at least four different vocal registers
existed, but that a definitive physiological and acoustic definition of all
register phenomena was not possible at that time (Hollien, 1985). The committee agreed that registers:
1.
involve a
series of consecutive fundamental frequencies that have the same perceived
timbre;
2. can be detected perceptually, and thus should be
recorded in the spectra of the different timbre groups;
3. are initiated by changes of laryngeal physiology
involving at least the internal muscles of the larynx.
The committee's
report questioned the scientific usefulness of the older names for registers
such as head and chest. The historic terms
attribute the identification of registers to areas of vibratory sensation in
singers. While vibratory sensations
definitely occur, they were not considered to be defining characteristics of registers. Defining characteristics are the physical and
acoustic events that give rise to the sensations.
While the
committee agreed that the sensations may be helpful in the education of singers
and speakers, they could not accept them as scientific evidence for defining
registers. The sensations themselves and their perceived
intensity vary widely between human beings, and replication of vibratory
sensations in groups of singers was nearly impossible to measure and study with
precision. Differences among perceived
vibratory sensations are the result of differences in: (1) anatomic structure and dimension among people,
(2) the nature of the physical coordinations used, (3) acoustic consequences in
body tissues, and (4) sensory perception abilities. The ability of the interoceptive and
proprioceptive sensory networks to bring vibratory sensations to conscious
awareness is variable. Interpretations
and verbal descriptions of their causes are subjective, and therefore, may be
inconsistent across human beings.
In order to
begin the process of register identification and definition, the committee
report identified four registers based on research at that time. In an attempt to reduce semantic confusion,
numbers were used to refer to the registers.
They were designated as #1, #2, #3 and #4 (Hollien, 1985). Based on information from the tradition of
singing pedagogy, a possible middle register between #2 and #3 was added and
designated #2A (e.g., Hollien & Schoenhard, 1983a,b). No scientific evidence for the existence of
this register was found by the committee at that time. The committee agreed that registers #2, #2A,
and #3 are the most frequently used in singing.
As voice science
and voice medicine became more prevalent, the National Institute on Deafness
and Other Communication Disorders (NIDCD), a component of the
As a result of
these developments, research into the phenomena of vocal registers has
increased over the past 25 years. In
addition to Harry Hollien, four voice scientists have consistently allied
themselves with various scientist and pedagogical colleagues to collaborate in
the study of vocal registers and associated voice qualities: Jo Estill, Donald Miller, Johann Sundberg,
and Ingo Titze (see references).
Within the vocal
pedagogy tradition—and among voice scientists—many questions have been raised
by: (1) the wide variety of register
concepts, terminologies, and practices, and (2) the variability of register
transition areas in the same voice
(Mörner, et al., 1964; Timberlake, 1990).
Eight explicit or implicit assumptions are imbedded in the current
jargon of vocal registers that are likely to be confusing to people who are
voice terminology novices:
1.
There
are speaking-voice
registers and singing-voice registers. An implicit assumption is that all human
beings have two voices, one for the “speaking voice”, and one for the “singing
voice”, and each “voice” has categorically different vocal registers.
2.
Chest
register is
associated with lower singing pitch range and a comparatively “thicker” voice
quality. An implicit assumption is that
it is activated by neuromuscular coordinations, or other phenomena, that occur
within the chest and thus produces perceivable vibration sensations therein.
3.
Head
register is
associated with higher singing pitch range and a comparatively “thinner” voice
quality. An implicit assumption is that
it is activated by neuromuscular coordinations, or other phenomena, that occur
within the head and thus produces perceivable vibration sensations therein.
4.
Falsetto
register is
associated with highest singing pitch range, or with all pitches produced above chest register, and a
comparatively “thinnest” (or “thinner”) voice quality. This concept has confusing implications. In Western cultures, it is strongly
associated with a female-like voice quality produced by males, but is a “false”
or “fake” voice that is of little or no practical use except in comedy. Vocal jargon novices may ask, “Do females
have a falsetto register?” and, “If ‘falsetto’ refers to the voice quality
that occurs in all pitches above chest register, how is it that at least two
categorically different voice qualities can be produced above chest
register? Does that not violate the
basic definition of a vocal register?”
5.
When
voices change from one register to another, unskilled vocalists typically
experience register breaks (abrupt transitions), but skilled vocalists
typically experience blended register transitions. Vocal jargon novices may ask, “What vocal
anatomy and physiology creates a ‘voice break’ in one person but not in another
person?”
6.
Middle
register is
associated with a middle singing pitch range and a voice quality that is a
“mixture” of chest and head (or falsetto)
registers. This concept also has
confusing implications. Vocal jargon
novices may ask, “How does one ‘mix’ two categorically different voice
qualities that, presumably, are produced by unique physiological
coordinations?” and, “If ‘falsetto’
refers to the voice quality that occurs in all pitches above chest register,
then how does this concept make sense?”
7.
A
lower and an upper passaggio pitch area are in all voices and they define the
lower and upper pitch range compass of middle register. Vocal jargon novices may ask, “How does this
concept make sense in the context of items 1 – 5 above?
8.
Each
register can be performed throughout the entire capable pitch range of all
singers, from lowest capable pitch to highest.
Vocal jargon novices may ask, “How does this concept make sense in the
context of all the above items?
What are vocal
registers, really? What anatomy and
physiology produce their acoustic phenomena?
How many registers are there?
What are the most accurate and helpful word labels for vocal registers? What happens physically and acoustically when
register events occur? Can the pitch
areas where register transitions occur be changed, or do they indicate
unchangeable, genetically inherited vocal characteristics? How is it that there can be so many different
register patterns in voices (e.g., strong lower-pitch-range registers and
weak/breathy upper-pitch-range registers, or vice versa, and register
transitions that occur around several different pitches)? How does “belting out a song” relate to
registers, and are there voice health issues involved in the use of belted
singing? How do registers and their
sound qualities relate to the musical styles of the world's cultures and
sub-cultures?
CONTEXT FOR A SCIENCE-BASED THEORY OF
VOCAL REGISTERS
An important
conceptual understanding that underlies this paper is that scientific
investigation is carried out by human beings and is, therefore, imperfect. Originally, the scientific method was
invented as a means of determining objective
reality and thus overcoming subjective
human bias, so that valid and reliable knowledge could be gathered. Totally “objective” scientific investigation
implies that the human beings who engage in scientific investigation are
entirely free from implicit assumptions and that they can disconnect the parts
of their brains that process feelings-emotions (biases) from those brain parts
that process perception and analytical conceptualization.
For instance, in
scientific investigations there is a possibility that human
investigators—inside or outside their conscious awareness—may orient research
procedures and findings so as to conform with previously held, emotionally
nuanced points of view. In addition,
technological instrumentation that is used to gather data for analysis may not
be sensitive enough to detect all of the phenomena that are relevant to a given
investigation, and sometimes, the instrumentation that could gather needed
information may not yet have been invented.
In spite of
these realities, the methods of science are still the best means we human
beings have yet devised to minimize
the influence of human bias. The “saving
grace” of scientific investigation is that, over time, human scientists will
question and reinvestigate previous findings and, based on a preponderance of
evidence, reconfigure theoretical explanations of “the nature of the world”.
The authors of
this paper do not claim status as “voice scientists”. We do claim to do the best we can to be
current with findings in the voice sciences, and to integrate that information
with our experience in helping people who are learning to sing and speak with
increasing skill and expressiveness. We
believe that such a background gives us credible grounds upon which to propose
a science-based theory of vocal register phenomena that we believe can resolve
historic conceptual, terminology, and practice discrepancies. In doing so, we hope to decrease doubts about
the credibility of the voice professions that occur among some of the people
who expect a high degree of consensus in voice-function knowledge among voice
scientists, speech pathologists, singing and speech teachers, choral
conductors, music educators, and theatre directors.
Some readers of
this paper are quite familiar with the scientific language of vocal anatomy,
physiology, and acoustics. Other readers
may be less familiar or unfamiliar with that language. This paper will attempt to present the theory
with both reader groups in mind. It also will assume that during the register
phenomena that we describe, (1) vocal anatomy and physiology are in a state of
health, and (2) the neuromuscular coordinations that enact basic breathflow,
phonation, and resonation are reasonably efficient. The authors acknowledge that some aspects of
this theory of vocal registers are not yet fully substantiated by scientific
research. If some of its provisions are
shown eventually to be inaccurate, that will be a learning moment, and that
learning will be celebrated because learning is what human beings do.
Two commonly
used linguistic nominalizations are: speaking voice and singing voice. The terms
denote concrete categorical differences between two “voices” in human beings,
but of course, human beings have one voice (one larynx and vocal tract), and
its neuromuscular coordinations produce all vocal phenomena, including speaking
and singing (details in Endnote 1). For
that reason, in this paper, there are no references to a so-called speaking voice and singing voice. Vocal
register phenomena, therefore, occur in all vocal sound-making, speaking, and
singing.
Using the
metaphor of a theatrical production, here is a review of the anatomy,
physiology, and acoustic processes that we regard as relevant to producing the
vocal phenomena that are referred to as vocal
registers.
The Producer is the genetic and epigenetic
expression that forms and maintains vocal anatomy and the neuropsychobiological
processes that activate and modulate vocal physiology.
The Playwright and the Technical and
Performance Director is the
central nervous system (CNS; brain and spinal cord). The CNS contains the vast neural networks,
and networks of networks ad infinitum,
that plan and enact the complex neuromuscular coordinations that produce all
overt and covert physical movements, including vocal phenomena such as vocal
registers. Learning new vocal abilities,
or altering already learned abilities, can only occur if relevant neural
networks are added to, or altered (for general reviews, see Fuster, 1997, 2003;
Holstege, et al., 1996; Huttenlocher, 1994; Thurman & Welch, 2000, Book I,
Chapters 3-9; Verdolini & Titze, in preparation).
The endocrine
and immune systems are Assistant
Directors that are interfaced with and modulate nearly all physical
functions, including those of the CNS.
Together, the Director and Assistant Directors coordinate all human
neuropsychobiological processing, including self-expression through symbolic
systems (languages, mathematics) and symbolic modes (music, dance, theatre,
painting, sculpture, architecture, and the like).
The Stage Crew is the peripheral nervous
system (PNS). It is made up of a somatic
division (cranial and spinal nerves) and an autonomic division (sympathetic,
parasympathetic, and enteric subdivisions).
The PNS has both sensory and motor nerves that are the interface between
the CNS and the external world, and between the CNS and internal bodily
processes. The motor functions of the
PNS are activated by integrative processing between sensory perception and
executive functions of the CNS.
The
playwright/director and crew have collaborated in a whole series of hit vocal
register dramatic and musical plays, such as:
A Streetcar Named Registers
A Funny Thing Happened on the Way to the
Registers
The Register Menagerie
Paint Your Registers
Joseph and the Amazing Technicolor
Registers
The Secret Register
How to Succeed in Registers without Really
Trying
Les Régisterable
The Leading Actors in these productions are
the primary laryngeal muscles that
induce the primary acoustic phenomena
of vocal registers. The role names of the leading actors refer
to the primary functions of the
“leading actor” muscles. (see Table 1)
1. Each of the
paired thyroarytenoid muscles (TA)
have two parts, a vocalis part
(thyrovocalis) and a muscularis part
(thyromuscularis). The thyrovocalis
parts extend for most of the length of the vocal folds and form their body or
core. The primary role of the TA muscles is to have a vocal fold shortening
influence within the synergistic functioning of all the internal larynx muscles
(especially in their interactions with the cricothyroid muscles). The
thyrovocalis parts appear to perform most of the shortening influence and both
parts appear to have a secondary adductory influence on the folds. For people who might feel averse toward the
use of anatomic terminology, or are too young or inexperienced to use it, the
vocal fold shortener muscles can be a
colloquial English term for the primary role of the thyroarytenoid muscles.
2. Each of the
paired cricothyroid muscles (CT)
have two parts, a more upright part (pars
recta) and a more oblique part (pars
obliqua). The most anterior ends of
the CT muscles are attached to the front of the cricoid cartilage and the
posterior ends are attached inside the lower lateral walls of the thyroid
cartilage. The primary role of the CT muscles is to have a vocal fold lengthening
influence within the synergistic functioning of all the internal larynx muscles
(especially in their interactions with the thyroarytenoid muscles). Their agonist-antagonist action with the
thyroarytenoid muscles creates a complex kind of “rocking” motion between the
cricoid and thyroid cartilages that alters the length of the folds. For people who might feel averse toward the
use of anatomic terminology, or are too young or inexperienced to use it, the
vocal fold lengthener muscles can be
a colloquial English term for the primary role of the cricothyroid muscles.
3. The spatial
location and configuration of the cover
tissues of the two vocal folds (official term: lamina propria) is altered by actions of all the internal larynx
muscles. The synergistic influence on
the folds’ cover tissues by the vocal fold shortener and lengthener muscles is
of greatest relevance to a science-based theory of vocal registers. The more the vocal folds are shortened, the
more thickened and lax the cover tissues become, and the more the vocal folds
are lengthened, the thinner and more taut the cover tissues become. These changes in the configuration of the
oscillating vocal folds influence the modes
of their oscillation and they change the characteristics of the voice source
spectrum.
The Major Supporting Actors are the
laryngeal muscles that induce a secondary
influence on the acoustic phenomena of vocal registers. The role
names of the major supporting actors refer to the secondary functions of these “supporting actor” muscles. (see Table
1)
1. The paired posterior cricoarytenoid muscles (PCA) are located at the rear area of
the larynx on the right and left sides.
They are attached to the cricoid and arytenoid cartilages in such a way
that they participate in abducting the arytenoid cartilages and thus opening
the vocal folds. Their primary role, therefore, is to have a
vocal fold opening influence within the synergistic functioning of all the
internal larynx muscles, especially in their interactions with the lateral
cricoarytenoid and the interarytenoid muscles.
Voice terminology novices may be more comfortable referring to these
muscles as the primary vocal fold opener
muscles.
2. The paired lateral cricoarytenoid muscles (LCA)
are located on the right and left sides of the larynx. They are attached to the cricoid and
arytenoid cartilages in such a way that they participate in adducting the vocal
processes of the arytenoid cartilages, and thus closing the vocal folds. Their primary
role, therefore, is to have a vocal fold closing influence within the
synergistic functioning of all the internal larynx muscles, especially in their
interactions with the posterior cricoarytenoid and the interarytenoid
muscles. Voice terminology novices may
be more comfortable referring to these muscles as one of the primary vocal fold
closer muscles.
Table
1
Summary of Internal Laryngeal Muscles
that Can Interact
to Produce Voice Source Spectra
Variations
that Can Be Perceived as Basic Voice
Quality Variations
|
Muscles |
Functions and
Influences on Voice Source Spectra |
|
Interarytenoids
(IA) |
·
A primary adductor of the cartilagenous portion of the
vocal folds |
|
Lateral
cricoarytenoids (LCA) |
·
A primary adductor of the membranous portion of the vocal
folds; ·
Agonist-antagonist interaction with IA and PCA to stabilize
vocal folds in many specific adductory positions |
|
Posterior
cricoarytenoids (PCA) |
·
Primary abductor of vocal folds; ·
Agonist-antagonist interaction with IA and LCA to
stabilize vocal folds in many specific adductory positions |
|
Thyroarytenoids
(TA) |
·
Primary shortener and thickener of the vocal folds ·
Secondary adductor of the vocal folds ·
Agonist-antagonist interaction with CT to stabilize the
length of the vocal folds in many non-specific and specific “settings” to
produce a wide range of F0s ·
Primary lengthener and shortener of the vocal folds when
not opposed by action of the CT |
|
Cricothyroids
(CT) |
·
Primary lengthener and thinner of the vocal folds ·
Agonist-antagonist interaction with TA to stabilize the
length of the vocal folds in many non-specific and specific “settings” to
produce a wide range of F0s ·
Primary lengthener and shortener of the vocal folds when
not opposed by action of the TA |
3. The singular interarytenoid muscle (IA) is located
at the posterior areas of the two arytenoid cartilages. It is attached to the arytenoid cartilages in
such a way that it participates in adducting the rear areas of the arytenoid
cartilages, and thus the cartilagenous portion of the vocal folds. Its primary
role, therefore, is to have a vocal fold closing influence within the
synergistic functioning of all the internal larynx muscles, especially in their
interactions with the posterior and the lateral cricoarytenoid muscles. Voice terminology novices may be more
comfortable referring to these muscles as one of the primary vocal fold closer muscles.
Plotline Settings for the “Register Plays”
PLOT SETTING 1—Auditory Perception and
Memory of Acoustic Phenomena. When bodymind auditory systems perceive a
series of sound events that share the same (or nearly the same) acoustic
characteristics, then those characteristics are correlated within a number of
interconnected auditory neural networks to become a larger neural network that
extends into both the parietal and frontal brain areas (see Fuster, 2003). As a result, a distinct perceptual category
is instantiated within the neural networks—referred to as memory.
For instance,
when a range of different fundamental frequencies (F0s) are produced
but prominent spectral characteristics remain nearly the same, then a perceptual
category of perceived voice quality is formed even though the F0s
have changed. But when a range of F0s
occurs and a different set of
spectral characteristics are produced, then a slightly different combination of
neural networks process those events and another distinct perceptual category
is formed in memory. Language labels are
not needed in order for such percepts to occur, but typically, such labels are
assigned by human beings.
PLOT SETTING 2—Vocal Fold
Oscillation and Its Two Primary Modes. The most widely known
theory of how mucosal waves are initiated and sustained is called the myoelastic-aerodynamic theory of vocal
fold vibration (Van Den Berg, 1958). The theory proposes that vocal fold
vibration (complex mucosal waving) occurs when:
1. the vocal fold surfaces are sufficiently compliant and elastic;
2. the vocal
folds are adducted enough to create a sufficiently narrow glottis; and
3. the
pressure-induced airflow force is great enough.
During each oscillation cycle of the adducted vocal folds,
the folds are in alternate closed and open phases. During the closed phase of each vocal fold cycle, subglottal air pressure very
rapidly builds up enough to displace the surface layers of vocal fold tissue
and trigger an open phase. At one time, the Bernoulli effect was thought
to bring the vocal folds back together for the next closed phase, but in 1988,
scientific reservations were expressed (Titze, 1988; see also Titze, 2000, pp.
109-111). While the Bernoulli effect is
incidentally present, its influence has been considerably overstated in prior
versions of the myoelastic-aerodynamic theory of vocal fold vibration. The greater influence over the return of the
vocal folds from open phase to closed is:
1. the
constraining elastic properties of the folds themselves which reverse their
opening motion back toward closure, and
2. “...the
synchrony between the driving (subglottal) pressure and (alterations in) tissue
velocity...” during mucosal waving cycles (Titze, 2000, p. 110; parenthetical
expressions added for clarity).
Finally, there are two primary
modes of vocal fold vibration. One mode
can be described as a repeated medial-to-lateral-to-medial-to-lateral (and so
on) oscillatory motion. The other mode
can be described as a repeated bottom-to-top waving oscillitory motion that
mostly occurs in the more surface tissues of the vocal folds. In most speaking and singing, the two modes
occur simultaneously.
PLOT SETTING 3—Contributions to Basic
Voice Qualities by Internal Larynx Muscles and the Vocal Tract. The
vocal fold closer and opener muscle groups primarily
(not exclusively) contribute one range of basic voice qualities (see Figure 1),
and the vocal fold shortener and lengthener muscles contribute to another range
of basic voice qualities (registers, see later). When vocal folds are incompletely adducted to
some degree, they oscillate to create vocal sound waves, but pressurized air
molecules also are flowing through the opening between the folds, thus
producing air turbulence noise. A
combination of vocal tone and air turbulence noise produces a breathy family of voice qualities.
|
Figure 1 Vocal Fold Closer Muscles’ Contributions to Basic Voice Qualities |
|||||
|
Breathflow “Energy” |
Muscle Energy |
||||
|
Whisper- |
Breathy |
Clear
& Richer Family |
Pressed-Edgy Family |
||
|
Noise |
Family |
|
Tense |
||
|
Family of |
|
P |
mf |
f |
Strained |
|
Sound |
|
firm |
richer |
richest |
Strident |
|
Qualities |
|
flutier |
mellow |
brassier |
Constricted |
|
|
|
|
warm |
|
Harsh |
When vocal folds
are adducted with a range of higher forces, and lung-air pressure is
correspondingly high, a pressed-edgy
family of voice qualities is produced.
Other terms used for this range of voice qualities are tense, tight, strained, strident, constricted, and harsh.
When vocal folds
are sufficiently adducted and “balanced” with appropriate lung-air pressure, a clear and richer family of voice qualities
can be produced. Because degrees of
adductory force are a major source of vocal fold amplitude/intensity during
speaking and singing, this family of voice qualities changes with perceived
vocal volume, but none of these voice qualities have breathy or pressed-edgy
characteristics. At softer volume
levels, a firm (not breathy) and flutier voice quality can be
perceived. At “middle-loud” volume
levels, a richer (more upper
partials) but mellow-warm voice
quality can be perceived. At “loud”
volume levels, a richest (even more
upper partials) and brassier voice
quality can be perceived.
When the vocal
tract changes its length and/or circumference dimensions, it has differential
effects on the pressures within the voice source sound spectra that are passing
through it (see Figure 2). Very
generally, when vocal tract dimensions are enlarged toward an extreme, lower
partials are amplified and upper partials tend to be damped, thus influencing
an over-full or over-dark family of voice qualities. Accordingly, when vocal tract dimensions are
diminished toward an extreme, higher partials are amplified and lower partials
are significantly damped, thus influencing an over-bright family of voice qualities. When vocal tract dimensions are optimally
configured, a relatively balanced complement of higher and lower partials pass
through and out of the vocal tract thus influencing a balanced resonance family of voice qualities that can range between
fuller and brighter.
|
Figure 2 Vocal Tract
Contributions to Basic Voice Qualities |
||
|
Overdark Family |
Balanced Resonance Family |
Overbright Family |
|
[“Ten-Foot
Giant” |
“Sweet Spots” |
“Bugs Bunny”] |
|
|
|
|
|
Throaty or
“Back” |
ç Fuller |
Narrow |
|
Sob-like |
Brighter è |
Squeezed |
|
Woofy |
|
Pinched |
|
Bottled-Up |
|
Piercing |
PLOT SETTING 4—Resonance Effects on Vocal
Fold Tissues and Internal Larynx Muscles. Above the larynx and
its vocal folds, there is a “tube” of sorts.
Its two ends, vocal folds and lips, can be open or closed. This tube is officially labeled as the vocal
tract because it is the tract through which vocal sound waves pass, and it can
vary its dimensions in numerous ways.
Very generally, two vocal tract cavities can open or narrow: (1) the
pharyngeal cavity (“throat part”) and
(2) the oral cavity (“mouth part”).
During speaking and singing, the vocal tract provides an open end (the
lips or teeth) from which vocal sound waves can radiate.
The trachea (“windpipe”) is a rounded tube that extends
downward from the larynx and its vocal folds.
Its general dimensions, especially length, increase in children as they
grow to their adult size. Dimensions
vary between adult male and female human beings, and its length and
circumference can be altered physiologically to a degree. In ordinary life circumstances, most of these
dimensional differences are comparatively small. Just like all tubes, of course, the trachea has a resonance frequency.
Vocal sound pressure waves are created by the oscillating
vocal folds and then radiate upward
through the vocal tract. Simultaneously,
sound pressure waves also are created that radiate downward into, and “bounce around” within, the trachea. Those sound pressure waves are “trapped”
inside the trachea, so during voicing, they are continually impacting on the
underside of the vocal folds. As the
fundamental frequency of those sound waves approaches and then matches the
resonance frequency of the trachea, the pressures within the reverberating
tracheal sound waves are increased (reinforced or amplified). In other words, those sound waves have
greater intensity, and the force of their impact on the underside of the vocal
folds is that much greater.
The increased pressures can produce an
interference with vocal fold oscillations where they are initiated, on the underside (trachea side) of the vocal
folds. The interference has been
referred to as acoustic loading of the
vocal folds. Acoustic loading has
been suggested as a significant influence on the passaggio effects that are related to vocal registers and their
transitions (Austin, 1992; Austin & Titze, 1997; Titze, 1983, 1984, 1988, 2000).
Even
when acoustic loading is not present, acoustic
overloading of the vocal folds
can occur. When the throat or mouth
areas of the vocal tract, or both, are sufficiently narrowed, the sound
pressure waves within the vocal tract are deflected and begin to reverberate
inside the vocal tract. These sound pressure
waves can interfere with vocal fold oscillations as they are passing over the topside (throat side) of the vocal
folds. Sound pressure wave impacts,
combined with the always-occurring tracheal sound pressure wave impacts, then
produce an acoustic overloading effect.
PLOT
SETTING 5—Activation of the Larynx by Neuromotor and Neurosensory
Processing. Two general types of motor functions occur in human beings: (1)
Reflexive or involuntary functions, and (2) learned or voluntary functions (see
Endnote 2 for background). Reflexive
vocal-motor functions (e.g., vocal startle sounds) are initiated from the
paired nucleus ambiguus areas of the brainstem's medulla oblongata and pass
through the peripheral nervous system's paired tenth cranial (vagus) nerves to
the relevant muscles (Hollien & Gould, 1990; Webster, 1995, pp.
282-298).
Deliberate or
learned vocal-motor functions are initiated in areas within the frontal
lobes of the two cerebral hemispheres, using sensory and/or memory input for
guidance (see Fuster, 1997, 2003).
Eventually, habitual learned
vocal-motor functions are triggered
within the motor areas of the frontal lobes, but nearly all of the actual motor
coordinations are enacted by
subcortical neural networks within the basal ganglia, cerebellum, and brainstem
(see Endnote 3 for learning background).
The left and right superior
laryngeal nerves (SLN) branch away from the paired vagus nerves to innervate the left and right sides of the
larynx (see Figure 4). The external branches of the SLN only supply motor innervation to the paired cricothyroid muscles, which
are the primary vocal fold lengtheners. The
internal branches of the SLN supply sensory reception only for the laryngeal mucosa that is immediately
above vocal fold level, and for some muscles of the larynx.
The left and right recurrent
laryngeal nerves (RLN) (see Figure 3) supply
motor innervation for all internal laryngeal muscles except the cricothyroids. That includes the thyroarytenoid muscles—the
primary vocal fold shorteners—and the adductory and abductory muscles that
close and open the vocal folds, respectively.
They also supply sensory reception
for the laryngeal mucosa that is immediately below the vocal fold level,
and for some muscles of the larynx.
|
Figure 3:
Illustration of key speech-voice motor areas in the central nervous
system and their connection to cranial nerve X (vagus) in the peripheral
nervous system. [From Professional Voice: The Science and Art of
Clinical Care, 2nd edition, by
R.T. Sataloff (Ed.) ©1997. Reprinted
with permission of Delmar, a division of Thomson Learning. FAX 800 730-2215.] |
|
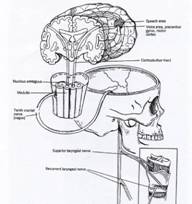
|
Figure
4: Illustration of the peripheral
motor and sensory innervation of the larynx.
[From Aronson, A. (1980). Clinical
Voice Disorders (2nd Ed.). |
|
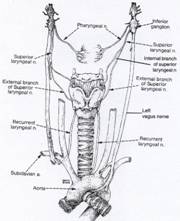
To
appreciate the capabilities of the larynx for speed and precision of its
neuromuscular coordinations, an understanding of its muscle fiber types and
related motor unit types is necessary (see Table 2 and Endnote 2 for
background). Normal genetic endowment
provides a greater percentage of Type S motor units and their Type I muscle
fibers in most of the external
laryngeal muscles. The internal laryngeal muscles have all
categories of the motor unit and muscle fiber types, including a significant
number of the S motor units and Type I muscle fibers, BUT Type FR and FInt motor units and Type IIA fibers are
predominant (see Table 2 and Endnote 1; also Bendiksen, et al., 1981; Claasen
& Werner, 1992; Cooper, et al., 1993; Titze, 2000a). The small internal laryngeal muscles are
estimated to have about 100 motor units per muscle. The capability, therefore, for high
variability in the motor unit recruitment patterns and action potential
frequencies is present in laryngeal muscles.
That means that laryngeal muscles are capable of a fairly wide range of slow-to-fast contractile speeds
and have the capability for: (1) extensive and vigorous use, (2) high
agility, (3) subtle, intricate adjustments, and (4) considerable resistance to
fatigue when they are activiated with optimum efficiency and are well
conditioned. In fact, muscles of the
larynx are regarded as having the second fastest contraction capability in the
whole body (Mårtensson & Skogland, 1964; eye muscles are fastest). That capability is related to survival
functions such as: (1) preservation and
facilitation of breathing, (2) high-speed closing of the airway to protect the
lungs, and (3) making loud sounds quickly to frighten predators (vocal startle
response).
Table
2
Muscle fiber types combined with motor unit types
(see Gordon & Patullo, 1993).
|
Muscle Fiber Type I: |
Slow-speed oxidative fibers (SO) that are highly resistant to fatigue, are smaller and unable to generate as much force as Type IIB fibers. |
|
Muscle Fiber Type IIa: |
Fast-speed oxidative and glycolytic fibers (FOG) that are moderately resistant to fatigue and intermediate in size and force generation. |
|
Muscle Fiber Type IIb: |
Fast-speed
glycolytic fibers (FG) that have low fatigue resistance, and tend to be
larger and capable of generating greater contractile force. |
|
Motor Unit Type S: |
Slow and
fatigue resistant (oxidative muscle fibers). |
|
Motor Unit Type FR: |
Fast and
fatigue resistant (oxidative and glycolytic muscle fibers). |
|
Motor Unit Type FInt: |
Fast and
fatigue intermediate (more glycolytic than oxidative fibers). |
|
Motor Unit Type FF: |
Fast and
fatigable (glycolytic muscle fibers). |
The realization of optimal high-speed response and fatigue
resistance in laryngeal muscles depends on the nature of their neural
input. Consistent with research in
neuromuscular physiology, a reasonable assumption can be made that when
laryngeal muscles are activated with reasonable frequency, but not very strenuously
over longer time periods, then presumably, Type S motor units will become
larger and their metabolic capacity changes so that the number of neural
impulses (action potentials) that they can generate increases, to some degree. Presumably, then, more protein will be added
to the Type I muscle fibers, thus increasing their size. More capillaries will be grown around those
fibers to supply bloodflow-delivered oxygen and nutrients in larger amounts.
When laryngeal muscles are engaged for shorter bursts of
strong, vigorous activity (such as shouting or sung pitches that are high and
loud), they develop fast speed capability by adding protein to the Type II
muscle fibers, thus increasing their size (bulk) and their capacity for
responding to more rapid motor unit neural impulses. Cellular changes also occur that enable
increased metabolic activity. When
speakers or singers must activate their laryngeal muscles with strength over
longer time periods, then protein is added to the more extensive Type IIA fibers
and cellular changes occur to increase both the speed of motor unit response
and resistance to fatigue. Well
conditioned laryngeal muscles are a fundamental requisite for skilled,
expressive speaking and singing (more below; see Saxon & Schneider, 1995;
Thurman & Welch, Book II, Chapter 15).
In the larynx, a voluntary control system has been
identified that initiates vocal fold closing and opening and lengthening and
shortening, and monitors their continuation so that adjustments can be made to
match desired vocal intentions (Larson, 1988; Strong & Vaughan, 1981;
Webster, 1995; Wyke, 1983a).
Neuromuscular motor networks are linked with sensory receptor networks
to form feedback loops to guide vocal coordinations toward fulfilling the
bodymind's intentions. Motor networks
signal selected muscles to contract in a particular sequences, speeds, and
intensities (for additional information on laryngeal capabilities, see Endnote
4).
Laryngeal motor networks are modified by innervation from
the sympathetic and parasympathetic divisions of the autonomic nervous system (ANS) (Basterra, et al., 1989). The ANS is prominently influenced by the
brain's limbic system, and together they sometimes are referred to as the emotional motor system. Feeling or emotional states, therefore,
affect vocal function (Graney & Flint, 1993; Holstege, et al., 1996;
Thurman & Welch, 2000, Book I, Chapters 7 and 8).
PLOT SETTING 6—Larynx Muscle and Vocal
Fold Tissue Conditioning. If we just consider the physical state of
larynx anatomy and physiology, then there are two conditioning categories that
need attention: (1) larynx muscles and
connective tissues, and (2) the vocal fold cover tissues.
A body’s skeletal muscles move a body’s skeleton. Larynx muscles are skeletal muscles that move
the skeletal parts of the larynx which, in turn, moves and “shapes” its
internal soft, non-muscle tissues. The
muscles of the larynx are made of the same types of anatomic material as limb
and torso muscles, and they respond to use in nearly the same ways. When the extent and vigor of neuromuscular
activation is increased or decreased, four characteristics change:
1. Strength. With increased use, the capacity for muscle
contraction intensity and force generation increases (sign of conditioning). With decreased use, the capacity for
contraction intensity and force generation decreases (sign of deconditioning).
Vocal Effects: Typically, reduced strength in the
closer-opener muscles results in some degree of reduced vocal volume and tonal clarity
capabilities, while increased strength results in some degree of increased
vocal volume and tonal clarity capabilities.
Reduced strength in the shortener-lengthener muscles results in some
degree of reduced pitch range and vocal register abilities, while increased
strength results in some degree of increased pitch range and vocal register
abilities.
2. Endurance. With increased use, higher contraction
intensity can be sustained for longer and longer periods of time before fatigue
sets in (sign of conditioning). With
decreased use, fatigue sets in earlier and earlier (sign of deconditioning).
Vocal
Effects: Reduced endurance in the larynx muscles
results in faster fatigue rates.
Increased endurance results in slower fatigue rates so that people can
engage in longer and longer and more vigorous voicing over time.
3. Precision, speed, and “smoothness” of neuromuscular coordinations.
With increased use, the precision, speed, and smoothness of
neuromuscular coordinations is increased (sign of conditioning). With decreased use, the precision, speed, and
smoothness of neuromuscular coordinations is diminished (sign of
deconditioning).
Vocal
Effects: With increased extent and vigor of voice use,
the motor nerves that activate larynx muscle coordinations are likely to
increase their size, their supply of neurotransmitter molecules at synapses,
their responsiveness to activation, their conduction velocity, and activation
precision. Thus, the capability for vocal
agility is increased (both pitch speed and pitch interval agility), as is the
capability for very subtle, fine-tuned motor abilities that relate to
increasingly accurate in-tune singing and blended register transitions.
With decreased extent and vigor of voice use, those properties decrease over
time.
4. Bulk.
With increased use over time, genes are activated to produce additional
constitutive protein within each of a muscle’s fibers (to increase contractile
properties), and thus, the size of
the whole muscle increases (hypertrophy; a sign of conditioning). With decreased use over time, an atrophic
process occurs and constitutive protein in each muscle fiber is gradually
reduced, and thus, the size of the whole muscle and its contractile properties
decrease (sign of deconditioning).
Vocal
Effects: As shortener muscle bulk increases, the cover tissues of the left fold are moved slightly
toward the right fold, and cover tissues of the right fold are moved slightly
toward the left fold. This change of
vocal fold status contributes an adductory advantage to the vocal folds that
helps optimize the efficiency of vibratory waving in the cover tissues during
speaking and singing. This adductory
advantage can result in greater clarity of vocal tone and increased vocal
volume capabilities.
As shortener muscle bulk decreases, cover tissues of the left
fold are moved slightly away from the right fold, and cover tissues of the
right fold are moved slightly away from the left fold. This change of vocal fold status contributes
an adductory disadvantage to the
vocal folds that can reduce the efficiency of vibratory waving of the cover
tissues during speaking and singing.
These effects may be observed visually during laryngeal videoendoscopy
as irregular configurations in the waving motions of the vocal fold cover
tissues. This adductory disadvantage can result in more air leakage
during voicing (decreased clarity of vocal tone) and in decreased vocal volume
capabilities.
When increased
conditioning moves the cover tissues of both vocal folds slightly closer to
each other, then a clear, non-breathy tone quality can be more readily produced
with less contraction intensity in the vocal fold closer muscles. In addition, greater strength in the closer
muscles enables them to produce more intense vocal fold closure to generate
greater vocal volume with optimal contraction intensity.
When decreased
conditioning moves the cover tissues of both vocal folds slightly away from
each other, then a breathy tone quality is likely unless the vocal folds are closed
with greater intensity than would be necessary with optimal conditioning. In addition, less strength in the closer
muscles, combined with the “debulking” of the shortener muscles, reduces vocal
volume capability.
The generic term for ligaments, tendons, and the like, is connective tissue. Typically, ligament tissue attaches muscles
to skeletal parts. Actually, that tissue
is diffused through the muscle to form its membranes, and so forth. The core function of connective tissue is to
hold a body’s skeleton together. Its
nature, therefore, is to shrink. Range
of skeletal motion at joints is diminished if connective tissue is continually
slackened, because it shrinks and then cannot allow the actual capable range of motion to occur. Appropriately stretching the connective
tissues in legs and arms, for example, increases the range of motion in
skeletal joints, and that increases limb movement capabilities for motor
skills.
Vocal
Effects: Larynx muscles
are attached to their skeletal parts by connective tissue (anterior and
posterior maculae flavae). If people
have only engaged in relatively quiet conversation during their lives, then the
connective tissue of their vocal fold shortener and lengthener muscles will
have shrunk to a degree. If they then
attempt to speak or sing what for them would be higher pitches, then the
lengthener muscles may have to work harder to stretch the folds to sufficient
length in order to produce the intended fundamental frequencies. The shrunken state of the connective tissue
also may prevent that person from singing as high as is actually possible. [Caution: “Stretching” the larynx’s connective tissues
can easily be “taken too far too soon” and result in muscle-ligament strain,
learned vocal inefficiency, and temporarily diminished vocal capability.]
When epithelial tissue endures relatively forceful impact
and/or shear stresses over time, the tissue adapts to the encountered
circumstances. Micro-level changes occur
within the tissues that increase restoration processes and an adaptation to
“demand” on the tissues. The immune
system is likely to produce inflammation in the affected tissues, and genes in
those tissues will activate to produce tissue changes that will make the tissues
“tougher” or more resilient. A good
analogy would be the reactions of ungloved “soft” hands when their surfaces
handle heavier and rougher-surfaced materials over time.
Vocal
Effects: With decreased
numbers of impact and shear stresses, the epithelium and other surface cover
tissues become “soft”, but with gradually increased numbers of impact and shear
stresses, the epithelium and other surface cover tissues undergo micro-level
changes that make them “tougher” or more resilient. That means that they can “take” more impact
and shear stresses for longer and longer periods of time before inflammation or
other defensive measures “kick in”. Hand
calluses and vocal fold nodules are similar tissue reactions to stronger impact
and shear stresses over time.
A CURRENT, SCIENCE-BASED
THEORY OF VOCAL REGISTERS:
THE VOCAL REGISTER STORYLINE
Titze (2000, p.
282) describes vocal registers as “...perceptually distinct regions of vocal quality that can be
maintained over some ranges of pitch and loudness” (italics added). Hollien (1974) described a vocal register as
“...a totally laryngeal event; it consists of a series or a range of
consecutive voice frequencies which can be produced with nearly identical phonatory quality...” (italics
added). He further stated that “...the
operational definition of a register must depend on supporting perceptual,
acoustic, physiologic, and aerodynamic evidence.”
During speaking
and singing, the primary
agonist-antagonist functions of the thyroarytenoid and cricothyroid muscles
result in changes of vocal fold fundamental frequencies (F0s), and
they do so by lengthening-shortening, thinning-thickening, and tautening-laxing
the folds. When these vocal fold
configuration changes occur, they alter the voice source spectra that are
introduced into the vocal tract. Voice
source spectra are then modified by the changing dimensions of the vocal tract,
but the vocal tract can only modify what the larynx introduces into it. Radiated spectra, therefore, will retain
various spectral characteristics that were introduced into the vocal tract when
the larynx and respiratory system created the original (voice source)
spectra. Listening brains then are
capable of perceiving voice quality changes that are originated at the larynx
level, and distinguishing them from voice quality changes that are induced of
changing vocal tract dimensions.
This theory of
vocal registers subscribes to the perspective that the voice qualities that are
referred to as vocal registers are brought into acoustic existence by varied
states of the oscillating vocal fold tissues.
Those varied states are preponderantly altered by variable coordinations
of the internal larynx muscles, but most predominantly by the thyroarytenoid
(shortener) and cricothyroid (lengthener) muscles.
We propose five
categories of shortener-lengthener muscle adjustments that produce five
perceivable categories of vocal register voice quality that we will describe in
anatomic, physiologic, and acoustic terms.
The voice quality categories are correlated with changes of vocal fold length-thickness-tautness and thus
with fundamental frequency (F0) and voice source spectra that are
perceived by listeners as pitch and voice quality or timbre.
We have selected
five word labels for the five vocal register voice quality categories according
to the following criteria. The word
labels must:
1. convey direct referential correlation
with at least one universal, measurable, and perceivable parameter of vocal
acoustics;
2. be “relatable” to vocal anatomy and
function as defined within the anatomical, physiological, and voice sciences;
and
3. be easy to assimilate into the colloquial
English of people who are not familiar with the technical jargon of the voice
professions.
The voice
register labels that we have selected are:
1. pulse
register
2. lower
register
3. upper
register
4.
falsetto
register for men, flute
register for women
5. whistle
register
Pulse Register
Pulse register
is produced when the cricothyroid muscles (lengtheners) are uncontracted so
that vocal fold length is determined solely by increases and decreases in the
contraction of the thyroarytenoids (shorteners). The vocal fold mucosa, therefore, is quite
short, thick, and lax. There is a
comparatively minimal range of subglottal air pressures and minimal adductory
force, resulting in a minimal aerodynamic flow between the vocal folds. Pulse register can be produced in both
speaking and singing.
One defining
sound characteristic of this register is a series of sound bursts with audible
gaps in between each burst. The recorded
waveforms show a series of “wave packets” with a temporal gap in between (see
Figure 5). Some vocalists can
intentionally shorten and lengthen the temporal gaps, mostly by subtle
increases and decreases of subglottal air pressure and aerodynamic flow and
slight alterations in the vocal fold shortener and closer muscles. When a vocalists’ vocal folds are thick
enough--by genetic endowment or by sufficient swelling--they are capable of
increasing the subglottal pressure and vocal fold adduction just enough to shorten
the temporal gaps to produce a range of very low-frequency sustained
tones. At the present time, speech-voice
professionals label the audible gap version of this register as vocal fry or fry. Presumably, this
perceived sound quality reminded some people of the sound of slow-frying
food. The CoMeT committee referred to
pulse register as Register #1.
The more a
pulsed F0 lowers past about 70-Hz, the more experienced voice judges
identify the continuing sound as a series of pulses with gaps (see Figure
5). The more a pulsed F0
raises above about 70-Hz, the more experienced voice judges identify continuing
sound as vocal tone rather than bursts and gaps. The 70-Hz mark is the average crossover
frequency between the perception of pulses and the perception of sustained
sound within the pulse register, but the crossover can occur anywhere between
60-Hz to 80-Hz (Hollien & Michel, 1968; Hollien, 1974, 1985; Titze, 2000,
pp. 283-288). Different vocal tract vowel
shapes can produce acoustic overloading of the vocal folds and thus interrupt
the continuation of pulsed sound (presented later and in Titze, 2000, pp. 286,
287). For instance, the vocal tract
opening and neck-throat ease of an /uh/ vowel is more conducive to continuation
of pulsed sound, whereas the tongue and lip narrowing of the vocal tract on an
/oo/ vowel is more likely to produce a degree of overloading. Modification of vocal tract vowel shape to
avoid the overloading, then, will be necessary for continuation of vocal
sound.
|
Figure 5: (a) is a graph that shows the percentage of
experienced voice judges that perceptually discriminated F0s that
were (1) continuous, sustained sound, versus (2) a series of sound pulses
with audible gaps in between. (b)
shows recorded waveforms of temporal gap pulses (vocal fry “wave packets”). [From
I.R. Titze, Principles of Voice
Production. Copyright © 2000, |
|
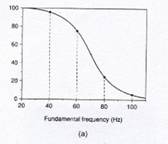
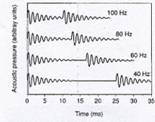
Pulse register
may be developed by some singers into an unusually low singing range. Fry also can be used as an initial pathfinder
(stepping-stone) to help some singers-in-training begin to develop their lower
register with physical efficiency. This
register is easier to produce when the vocal folds are swollen, so singers with
a history of fairly frequent tobacco smoking and alcohol drinking have much
greater chance of developing their pulse register coordination.
Some Russian and
Eastern European male classical music singers are well known for developing
this register and have become contrabass singers (German: Strohbass = straw bass, having a voice quality that suggests the
sounds that are made when straw is crushed).
In the choral singing of those cultures, contrabasses sometimes sing the
bass part one octave lower than the written notation, contributing to a
characteristically thick and dark tonal quality.
Members of some
Asian cultures use pulse register in chanting—Tibetan monks, for instance. Some cultures have developed highly skilled
“mouth” or “throat singing” that uses a sustained, very firm low-pitched, pulse
register drone to produce an array of overtones. The singers then shape their vocal tracts in
special ways to amplify harmonic regions so prominently that melodic contours
and other acoustic effects can be produced.
Lower Register
Lower register
voice qualities are produced when both the thyroarytenoid and the cricothyroid
muscles are simultaneously contracted (primary shorteners and lengtheners,
respectively), but the thyroarytenoids are more prominently contracted than the
cricothyroids (Hirano, et al., 1970; Vennard, et al., 1970a,b; Titze, 2000;
Titze, et al., 1989). Various
agonist-antagonist contractions of the two muscles result in a range of
stabilizations in vocal fold length, thickness, and tautness. The prominence of contraction by the
thyroarytenoid muscles results in generally shorter, thicker, more lax vocal
fold cover tissues and a lower range of F0s.
When compared to
the essential quality of upper register, the essential quality of lower
register can be described as thicker
and more full-bodied. That voice quality would be reflected in its
voice source spectra, with the lower partials having greater intensity when
compared to the lower partials of upper register voice source spectra. At the present time, various voice
professionals have labeled this register as chest
register, modal register, or heavy mechanism. The CoMeT committee referred to this register
as Register #2. We recommend the term lower register to reflect the pitch-dependent nature of this
register's laryngeal coordinations, and to eliminate the implicit assumption
that its activation occurs in the chest.
The increased
intensity in the lower partials of the voice source spectra is produced when a
greater mass of vocal fold tissue is involved in vocal fold oscillation (as
compared to thinner vocal fold tissue mass when upper register qualities are
produced). Greater tissue mass is
produced when the vocal folds are shorter and thicker, and thus, adduction of
both the superior and inferior areas of the vocal folds occurs (see Figure 6a;
Hirano, et al., 1970; Vennard, et al., 1970a,b; Titze, et al., 1989; Titze,
2000; Vilkman, et al., 1995). The
thyroarytenoid muscle, including the muscularis portion, bulges the portion of
the vocal folds that is below the level of the arytenoid cartilage's vocal
processes.
The vocal
ligaments also are more lax and can participate in vocal fold oscillation, and
the vocalis portion of the TA muscle undulates as well (Titze, 2000). The vocal fold ligament and the thyrovocalis
muscle tissues vibrate with much less amplitude, however, than the outer,
superficial layer of the cover because of their greater structural stiffness.
Typically, that means that during vocal fold oscillating there is:
1.
a
larger bottom-to-top contact area of the oscillating surface tissues; and
2. greater depth of tissue movement.
These
characteristics of vocal fold function result in longer closed phase times,
that is, the closed quotient (CQ) of each vocal fold oscillation is nearly
always above 50% of the total of each single oscillation cycle (0.5). These functions are observable in
electroglottographic (EGG) recordings.
The EGG recording in Figure 6b shows the greater CQ. The EGG for lower register shows a broader
peak than the one for upper register.
The “knee” in the lower register EGG waveform reflects the greater
contact time that is produced by the bulging of the vocal folds below the level
of the arytenoid vocal processes (Alipour & Scherer, 2000; Titze, 1990,
2000).
Lower Register Larynx Coordinations
Interfaced
with Vocal Fold Adduction Coordinations
In lower register, the TA muscles are
more prominently contracted than the CT muscles, thus:
1. the vocal folds are generally shorter so that a lower range
of F0s
is produced (when compared to upper register);
2. the folds are thicker and both the superior and the
inferior areas of the membranous portions are adducted, contributing to longer
closing phases in each oscillation cycle and greater depth of oscillation into
the membranous vocal fold tissues;
3. greater “bulging” of the TA muscle produces a secondary
adductory gesture, contributing to longer closing phases in each oscillation
cycle.
IN LOWER REGISTER WITH
INCOMPLETE VOCAL FOLD ADDUCTION, mucosal waving occurs to produce voice
source spectra, but air-turbulence noise is produced simultaneously when
subglottic airflow passes between the narrowed vocal fold opening. The vocal folds can create varying degrees of
incomplete adduction, with varying degrees of air-turbulence noise.
Because prominent TA contraction has thickened the vocal
folds and added a slight adductory gesture, the depth and inferior-to-superior
amounts of cover tissue involved in mucosal waving is greater when compared to
upper register with incomplete adduction.
Also, the closing phases last longer than the opening phases.
Intensity in the voice source spectra is increased by
increasing the amplitude of mucosal waving.
That is accomplished by increasing the amount of subglottic pressure and
aerodynamic flow. When subglottic
pressure is increased, however, the TA and CT muscles must subtly increase
their co-contraction in order to prevent a rise in intended F0 (singing sharp).
Intensity in the voice source spectra is decreased by decreasing the
amplitude of mucosal waving, and that is accomplished by decreasing the amount
of subglottic pressure and aerodynamic flow.
With decreases in subglottic pressure, the TA and CT muscles must subtly
decrease their co-contraction in order to prevent a lowering of intended F0 (singing flat).
Incomplete vocal fold adduction produces: (1) fewer upper
voice source spectrum partials, and (2) comparatively prominent intensities in
the F0s
and lowest harmonics of voice source spectra.
The result is relatively steep spectral slopes in which partial
amplitude reduces by at least 18-dB per doubling of the F0. In turn, these
conditions produce typical contributions to perceived voice quality which may
be described as airy and breathy but thicker and more full-bodied
when compared to upper register with incomplete adduction.
IN LOWER REGISTER WITH
INTENSE (PRESSED) VOCAL FOLD ADDUCTION, the degree of closure force creates
an impediment to transglottal airflow in such a way that the potential amplitude
of the vocal fold oscillations is inhibited.
The degree of intensity in the adductory force closes the glottal space
“too much” and prevents the vocal fold tissues from oscillating with optimum
amplitude. As a result, perceived vocal
volume is then less than what is possible and perceived voice quality may be
described as being pressed, tense, tight, constricted, or strained.
In addition, the excessive adductory force causes the
oscillating vocal folds to collide “too soon” and initiate a kind of
hyper-oscillatory mode of vibration in the vocal folds that produces
subharmonics. In other words,
“irregular” harmonics are created in the voice source spectra that are in between the regular harmonics. The additional harmonics create harmonic
dissonances that contribute a perception of voice quality that can be described
as edgy, buzzy, strident, or harsh.
The excessive intensity of adductory force tends to suppress
acoustic energy in the F0 and lower harmonics, but tends to produce a greater number
of higher partials in the voice source spectra, along with subharmonic partials
between them. The comparative
suppression of F0 and lower harmonic intensities results in more level
spectral slopes in which partial amplitude reduces by about 6-dB or less per
doubling of the F0. These conditions
add a brightness component to perceived voice quality that may be amplified by
a narrowed ary-epiglottic sphincter (epilarynx).
Because prominent TA contraction has thickened the vocal
folds and added a slight adductory gesture to them, however, the depth and
inferior-to-superior amounts of cover tissue involved in mucosal oscillation is
greater, when compared to the same conditions in upper register, and the
closing phase is longest of all. As a
result, the pressed and edgy perceived qualities can be
described as having a more full-bodied
or thicker component when compared to
the pressed and edgy qualities in upper register.
IN LOWER REGISTER, OPTIMAL
VOCAL FOLD ADDUCTION always produces vocal fold impacts (“collisions”)
during vocal fold oscillation, but allows enough glottal space for the mucosa
to achieve optimal amplitude and impact intensity. Subglottal air pressure and aerodynamic force
are increased when vocal volume is increased, and those conditions increase the
amplitude of mucosal waving. In order to
avoid pressed phonation, the agonist-antagonist action of the lateral
cricoarytenoid (LCA) and the posterior cricoarytenoid (PCA) muscles adjust the
vocal processes of the arytenoid cartilages toward a slightly more open
position to provide a slightly increased glottal space. That adjustment accommodates the increased
amplitude in vocal fold oscillations (the membranous portions of the two vocal
folds are extended medially into the glottal space), so that the folds impact
just when they need to for optimum impact (not “too soon”). The degree of the LCA-PCA adjustment depends
on the degree of desired vocal volume.
Because prominent TA contraction has thickened the vocal folds and added
a slight adductory gesture, the depth and inferior-to-superior amounts of cover
tissue involved in mucosal waving is greater when compared to upper register,
and the closing phase lasts longer than the opening phase.
These conditions contribute to an optimum number of higher
voice source spectrum partials, optimum intensity in the F0 and lower harmonics, resulting in a
spectral slope in which partial amplitude reduces by about 12-dB per doubling
of the F0. These acoustic characteristics produce
typical contributions to perceived voice quality which may be described as lighter-flutier, richer-warm/mellow, richest-brassier,
but thicker, more full-bodied, and heavier
when compared to upper register.
Upper Register
Upper register voice qualities are produced when both
the thyroarytenoid (primary shortener) and the cricothyroid (primary
lengthener) muscles are simultaneously contracted, but the cricothyroids are
more prominently contracted than the thyroarytenoids (Titze, 2000). Various agonist-antagonist contractions of the
two muscles result in a range of stabilizations in vocal fold length,
thickness, and tautness. The prominence
of contraction by the cricothyroid muscles results in generally longer,
thinner, more taut vocal fold cover tissues and a higher range of F0s (Hirano, Ohala, & Vennard, 1970;
Shipp & McGlone, 1971; Titze, 2000).
|
Figure 6: (A)
is a drawing that compares a cross-section of a right vocal fold configured
for upper register (left side) and for lower register (right side). |
Figure 6A Figure 6B |
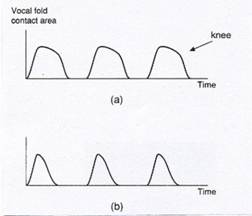
The essential
quality of this register, when compared to the essential quality of lower
register, can be described as thinner and lighter. The greater intensity that is present in the
lower partials of the lower register would no longer be present, and upper
register voice source spectra would reflect that loss. Typically, all of the partials in upper
register voice source spectra would have less overall intensity when compared
to lower register voice source spectra.
Proportionately, the partials nearest the F0
are the most intense. These partial intensity differences create
the distinct perceptual category described above.
The decreased
intensity in the voice source partials is produced when comparatively thinner
mass of vocal fold tissue is involved in vocal fold oscillation (as compared to
the thicker vocal fold tissue mass when lower register qualities are
produced). Thinner tissue mass is
produced when the vocal folds are lengthened and thinned, and thus, adduction
only of the superior portion of the vocal folds occurs (see Figure 6A). The vocal ligaments bear the vocal folds'
passive stretch tension so that only the lengthened and thinned epithelium and
superficial layer of the lamina propria can participate in oscillatory motion
(Titze, 2000; Vilkman, et al., 1995).
Typically, that means that during vocal fold oscillation there is:
1. a smaller
bottom-to-top contact area of the oscillating surface tissues; and
2. more shallow
depth of tissue movement.
These
characteristics of vocal fold function result in longer open phase times, that
is, the open quotient (OQ) of each vocal fold oscillation is nearly always
above 0.5. Some trained singers are able
to produce this register with a CQ that is slightly above 0.5 (Howard, et al.,
1995), presumably because they have the ability to strongly adduct their vocal
folds. These functions are observable in
electroglottographic (EGG) recordings.
The EGG for upper register shows a narrower peak than the one for lower
register, reflecting the fact that only the more superior area of the
bottom-to-top vocal fold mucosa is in contact (see Figure 6A and B).
At the present
time, various voice professionals label this register as head, falsetto, loft, or light mechanism. The CoMeT
committee referred to this register as Register
#3. In pre-scientific vocal pedagogy,
this register is labeled head by
some, and falsetto by others. Still others use falsetto for register #3 and head
for register #4. These varied uses have
produced considerable semantic confusion, especially among English-speaking
people. We recommend the term upper register to reflect the
pitch-dependent nature of this register's laryngeal coordinations, and to
eliminate the implicit assumption that its activation occurs in the head.
Upper Register Larynx Coordinations
Interfaced with Vocal Fold Adduction Coordinations
In upper
register, the CT muscles are more prominently contracted than the TA muscles,
thus:
1.
the vocal folds are generally longer so that a higher range of F0s is produced (when compared to lower
register);
2.
the folds are “stretched” thinner and only the superior areas of the membranous
portions are adducted, contributing to longer opening phases in each
oscillation cycle and a shallower depth of oscillation in the vocal fold cover
tissues;
3.
comparatively less “bulging” of the contracted TA muscles produces
comparatively less extensive secondary adductory gestures, thus contributing to
shorter closing phases in each oscillation cycle when compared to lower
register.
IN UPPER REGISTER WITH INCOMPLETE VOCAL FOLD ADDUCTION, mucosal waving occurs to
produce voice source spectra, but air-turbulence noise is produced
simultaneously when subglottic airflow passes between the narrowed vocal fold
opening. The vocal folds can create
varying degrees of incomplete adduction, creating varying degrees of
air-turbulence noise. Because prominent
CT contraction has thinned the vocal folds, the depth and inferior-to-superior
amounts of cover tissue involved in mucosal waving is less when compared to
lower register with incomplete adduction.
Intensity in the voice source spectra is
increased by increasing the amplitude of mucosal waving, and that is
accomplished by increasing the amount of subglottic pressure and aerodynamic
flow. When subglottic pressure is increased,
however, the CT and TA muscles must subtly increase their co-contraction in
order to prevent a rise in intended F0 (singing sharp).
Intensity in the voice source spectra is decreased by decreasing the
amplitude of mucosal waving, and that is accomplished by decreasing the amount
of subglottic pressure and aerodynamic flow.
With decreases in subglottic pressure, the CT and TA muscles must subtly
decrease their co-contraction in order to prevent a lowering of intended F0 (singing flat).
These conditions produce fewer upper
voice source spectrum partials, comparatively prominent intensities in the F0s and lower voice source harmonics,
resulting in relatively steep spectral slopes in which partial amplitude
reduces by about 18-dB per doubling of the F0 or more.
In turn, these conditions produce typical contributions to perceived
voice quality which may be described as airy
and breathy but lighter and thinner when
compared to lower register.
IN UPPER REGISTER WITH INTENSE (PRESSED) VOCAL FOLD ADDUCTION, the degree of closure
force creates an impediment to transglottal airflow in such a way that the
potential amplitude of vocal fold oscillations is inhibited. The degree of intensity in the adductory
force narrows the glottal space “too much” and prevents the vocal fold tissues
from oscillating with optimum amplitude.
As a result, perceived vocal volume is then less than what is possible,
and perceived voice quality may be described as being pressed, tense, tight, constricted, or strained.
In addition, the excessive adductory
force causes the oscillating vocal folds to collide “too soon” and initiate a
kind of hyper-oscillatory mode of vibration in the vocal folds that produces
subharmonics. In other words,
“irregular” harmonics are created in the voice source spectra that are in between the regular harmonics. The additional harmonics create harmonic
dissonances that contribute a perception of voice quality that can be described
as edgy, buzzy, strident, or harsh.
The excessive intensity of adductory
force tends to suppress acoustic energy in the F0 and lower harmonics, but tends to
produce a greater number of higher partials in the voice source spectra, along
with subharmonic partials between them.
The comparative suppression of F0 and lower harmonic intensities results
in more level spectral slopes in which partial amplitude reduces by about 6-dB
or less per doubling of the F0.
These conditions add a brightness
component to perceived voice quality that may be amplified by a narrowed
ary-epiglottic sphincter (epilarynx) and a shortened pharynx (elevation of
larynx).
Because prominent CT contraction has
thinned the vocal folds, there is less of the TA adductory gesture. The depth and inferior-to-superior amounts of
cover tissue involved in mucosal oscillation is less, when compared to lower
register, and the opening phase is nearly always longer than the closing
phase. As a result, the pressed and edgy perceived qualities can be described as having a thinner and lighter component when compared to the pressed and edgy
qualities in lower register.
IN UPPER REGISTER, OPTIMAL VOCAL FOLD ADDUCTION always produces vocal fold impacts
(“collisions”) during vocal fold oscillation, but allows enough glottal space
for the mucosa to achieve optimal amplitude and impact intensity. Intensity in the voice source spectra is increased by increasing the amplitude of
mucosal oscillation. To do that, the
vocal folds are adducted more firmly and that necessitates increased subglottic
pressure and aerodynamic flow in order to maintain oscillation. When subglottic pressure is increased,
however, the CT and TA muscles must subtly increase their already intense
co-contraction in order to prevent a rise in intended F0 (singing sharp).
Intensity in the voice source spectra is decreased by decreasing the amplitude of mucosal waving. To do that, the vocal folds are adducted less
firmly and that necessitates decreased subglottic pressure and aerodynamic
flow. With a decrease in subglottic
pressure, the CT and TA muscles must subtly decrease their co-contraction in
order to prevent a lowering of intended F0 (singing flat).
In order to avoid pressed-edgy voice
qualities during vocal volume increases, the agonist-antagonist action of the
lateral cricoarytenoid (LCA), the interarytenoid (IA), and the posterior cricoarytenoid
(PCA) muscles adjust the vocal processes of the arytenoid cartilages toward a
slightly more open position to provide a slightly increased glottal space. The degree of LCA-IA-PCA adjustment depends
on the degree of desired vocal volume.
Because prominent CT contraction has thinned the vocal folds, the depth
and inferior-to-superior amounts of cover tissue involved in mucosal waving is
less when compared to optimal adduction in lower register.
These conditions contribute to an optimum
number of higher voice source spectrum partials, optimum intensity in the F0 and lower harmonics, resulting in a
spectral slope in which the amplitude of partials reduces by about 12-dB per
doubling of the F0. These acoustic
characteristics produce typical contributions to perceived voice quality which
may be described as lighter-flutier, richer-warm/mellow, richest-brassier, but thinner
and lighter when compared to
lower register.
Falsetto Register (males)
and Flute Register (females)
Among speakers
of colloquial English, there is no confusion about the meaning of the term falsetto voice. It refers to a voice
quality that adult males can produce within the female pitch range and is
female-like in quality. Because of this
near-universal identification, labeling this register with any other term would
be confusing to a quite large majority of English speakers. Likewise, to use the same term for the same
laryngeal function in females also would be confusing. The term flute
was selected as the term for this register in females because its essential
quality resembles the tone quality of the flute instrument. It is borrowed from Hollien (1974).
Although there
is great variability among individual singers, and there are significant
differences of anatomical dimension between male and female vocal structures,
we propose that the basic biomechanics of falsetto/flute register are
essentially the same in both males and females.
The thyroarytenoid muscles (primary shortening influence) release
completely so that vocal fold length is determined entirely by action of the
cricothyroids (primary lengtheners) (Ardran & Wulstan, 1967; Titze, 2000;
Welch, et al., 1988). In addition, the
CT muscles are assisted by some of the external larynx muscles at the highest
and lowest F0s
(Vilkman, et al., 1995). Zero
contraction of the thyroarytenoids removes all of its shortening, thickening,
and laxing influences on the vocal fold cover tissues. In male falsetto, for instance, a typical
lower pitch range appears to be about E3 to C4.
In that pitch range, the vocal folds are as short and thick and lax as
they can be without activation of the thyroarytenoid muscles. Also, without the adductory gesture that is
provided by thyroarytenoid contraction, the membranous portion of the vocal
folds are likely to be separated (“bowed” configuration) and thus a breathy
quality is likely to be perceived.
As F0 is increased in the falsetto/flute
register coordination, optimal action by the primary adductor muscles
(closers), combined with the “stretching” (lengthening) action of the
cricothyroids, results in complete adduction and a fairly wide intensity
range. In this register's higher pitch
range, the vocal fold cover tissues are in their longest, thinnest, and most
taut range of configurations. In this
state, the ligament layers of the lamina propria bear even more of the passive
stretch tension than they exerted in upper register, and vocal fold oscillation
occurs only in the epithelium and the superficial layer of the lamina
propria. Typically, that means that
there is:
1.
a
thin vertical tissue mass that creates a thin bottom-to-top contact area for
the surface tissues;
2. minimal depth of oscillation in the vocal
fold cover tissues.
At the present
time, various voice professionals refer to this register as flute, whistle, or flageolet in
females and falsetto or pure falsetto in males. The CoMeT committee referred to the
falsetto/flute register as Register
#4. The essential quality of
falsetto/flute register, when compared to the essential qualities of the lower
and upper registers, can be described as lightest
and thinnest, although an optimally
longer vocal tract and expanded pharyngeal cavity can add “fullness” or a
“darker color” to the quality that is contributed by the vocal fold voice
source (Shipp, et al., 1988). The voice
source's contribution to perceived voice quality is reflected in its voice
source spectra. Typically, this register
coordination produces the fewest number of partials compared to the other
registers. The F0 is the most prominent partial, and the
overtones that are present have comparatively minimal intensity (Titze, 2000;
Walker, 1988). These voice source
spectrum characteristics resemble those produced when flutes are played in the
same F0
range, and result in a distinct perceptual category that identifies this
register.
When
falsetto/flute register coordination is used in its lowest F0 range, or when the laryngeal adductor
muscles are underconditioned, then the longest open phase times are produced
compared to upper and lower registers.
The OQ of each vocal fold oscillation is nearly always above about
0.7. These functions are observable in
electroglottographic (EGG) recordings.
The EGG for falsetto/flute register shows a narrower peak compared to
the one for upper and lower registers, reflecting the fact that a small amount
of the superior area of the vocal fold mucosa is waving. On the other hand, when falsetto/flute
register coordination is used in its middle to highest F0 range, or when the laryngeal adductor
muscles are well conditioned, the open phase times approach 0.5. In some professional falsettists and
countertenors, open phase falls below 0.5 (Shipp, et al., 1988; Welch, et al.,
1988, 1989).
Male falsetto
quality has been referred to with the value-laden term “effeminate” (Fuchs,
1963; Miller, 1977) and with such terms as “unnatural”, “artificial”, and a
“trick voice” that can only be performed at the pianissimo dynamic (Allen, 1935; Emile-Behnke, 1945). Its longer-term use has even been associated
with impaired vocal health (Miller, 1986, p. 122; Procter, 1980, p. 129),
although no evidence has been produced to verify such a claim. Videostroboscopic and electroglottographic
studies of professional male falsettists and countertenors have invalidated all
of the previous presumptions (Lindestad & Södersten, 1988; Welch, et al.,
1988, 1989). Many professional
countertenors use a form of upper register when they include a minimal degree
of thyroarytenoid contraction along with the degree of cricothyroid
contraction, a very fine-tuned neuromotor skill. The perceived quality is sometimes described
as upper register with a high percentage of falsetto quality “mixed in”, or as
falsetto register with a small amount of upper register “mixed in” (see Howard,
et al., 2001).
Falsetto/Flute Register Larynx Coordinations
Interfaced with Vocal Fold Adduction
Coordinations
In falsetto/flute register, the TA
muscles are not contracted at all. The
CT muscles perform both the primary lengthening and shortening of the vocal
folds, thus:
1. the vocal folds have their greatest range of elongation capability because there is
no antagonist resistance from the TA muscles;
2. the highest capable range of F0s is produced by all voices that have
normal anatomy and physiology, although whistle register coordination can
produce even higher F0s in the people who are capable of producing it;
3. vocal fold shortening is realized by reducing
the contraction intensity of the CT muscles so that the folds gradually become
slackened;
4. the vocal fold cover tissues have their
greatest range of thinning capability
[An absence of bulging by the TA muscles removes their thickening gesture, so
that only a tiny portion the superior areas of the membranous portions are
adducted. These factors contribute to a
range of longer opening phases in each oscillation cycle and a shallowest depth
of oscillating vocal fold cover tissue.];
5. absence of TA muscle “bulging” also removes their secondary
adductory gesture [Complete vocal fold closure is most easily accomplished when
the folds are stretched longer and are more taut. Typically, when singing or speaking in the
lower pitch range of this register, the absence of TA bulging and of lengthened
tautness produces a slackening of the folds, and their medial surfaces become
separated so that they take on a “bowed” configuration. That results in a breathy quality and a range of shorter closing phases in each
oscillation cycle when compared to upper register.].
IN FALSETTO/FLUTE REGISTER WITH
INCOMPLETE VOCAL FOLD ADDUCTION, mucosal waving occurs to produce voice
source spectra, but air-turbulence noise is produced simultaneously when
subglottic airflow passes between the open glottal area between the vocal
folds. The vocal folds can create
varying degrees of incomplete adduction with varying degrees of air-turbulence
noise. Because CT-only contraction has
thinned the vocal folds even more than in upper register, the depth and
inferior-to-superior amounts of cover tissue involved in vocal fold oscillation
is shallowest, when compared to upper register with incomplete adduction.
Intensity in the voice source spectra is
increased by increasing the amplitude of mucosal waving, and with incomplete
adduction, that is accomplished by increasing the amount of subglottic pressure
and aerodynamic flow. When singing in
the lower F0
range of this register, with slackened folds and breathy quality, the
subglottic air pressure must be comparatively minimal in order to avoid singing
sharp. The lower F0 range in this register, therefore, has a
quite limited intensity range. When
singing in the upper F0 range of this register, the vocal folds are progressively
“stretched” and tautened so that their vibratory edges are brought into closer
proximity and the adductor-abductor muscles can engage to maintain an
incomplete adduction. Under these
conditions, the vocal folds can provide greater “resistance” to subglottic air
pressure and the vocal intensity range capability is greater.
The above conditions produce fewer
partials in the voice source spectrum when compared to the upper register, and
there is a proportionately more prominent intensity in the F0.
The spectral slopes are comparatively steep and the amplitude of
partials reduces by at least 18-dB per doubling of the F0.
In turn, these conditions produce typical contributions to perceived
voice quality which may be described as airy
and breathy but thinnest and lightest
when compared to upper and lower registers.
IN FALSETTO/FLUTE REGISTER WITH
INTENSE (PRESSED) VOCAL FOLD ADDUCTION, the degree of closure force creates
an impediment to transglottal airflow in such a way that the potential
amplitude of the vocal fold oscillations is inhibited. The degree of intensity in the adductory
force narrows the glottal space “too much” and prevents the vocal fold tissues
from oscillating with optimum amplitude.
As a result, perceived vocal volume is then less than what is possible
and perceived voice quality may be described as being pressed, tense, tight, constricted, or strained.
In addition, the excessive adductory
force causes the oscillating vocal folds to collide “too soon” and initiate a
kind of hyper-oscillatory mode of vibration in the vocal folds that produces
subharmonics. In other words,
“irregular” harmonics are created in the voice source spectra that are in between the regular harmonics. The additional harmonics create harmonic
dissonances that contribute a perception of voice quality that can be described
as edgy, buzzy, strident, or harsh.
Pressed adduction and pressed-edgy
quality are not possible in the lower F0 range of this register coordination
because of the shorter, slackened state of the folds (absence of TA
contraction).
The excessive intensity of adductory
force tends to suppress acoustic energy in the relatively few partials that are
produced by this register coordination. but tends to produce a greater number
of higher partials in the voice source spectra, along with subharmonic partials
between them. The spectral slopes are
more level than those that are produced by optimal adduction. On average, partial amplitude reduces by at
least 6-dB per doubling of the F0, perhaps more. These
conditions add a brightness component
to perceived voice quality that may be amplified by a considerably narrowed
ary-epiglottic sphincter (epilarynx) and shortened pharynx (elevation of
larynx). The depth and
inferior-to-superior amounts of cover tissue involved in vocal fold oscillation
is quite minimal, when compared to upper register. The opening-to-closing phase ratio can
vary. Only some well conditioned
barbershop singers and professional countertenors and falsettists have produced
a closed phase that lasts longer than open phase (Penrose, et al., 2001).
IN FALSETTO/FLUTE REGISTER WITH
OPTIMAL VOCAL FOLD ADDUCTION, the slackened state of the vocal folds in the
lower F0
range prevents contact during mucosal oscillation, and breathy phonation is inevitable.
In the upper F0 range, the longer, more taut vocal fold cover tissues enable
complete adduction and clear voice
quality (no breathiness). Vocal volume
is increased when subglottal air
pressure and aerodynamic flow are increased.
Presumably, these conditions increase the amplitude of vocal fold cover
tissue oscillation. As a result, the
mucosal tissues “spread” medially into the glottal space to a degree. The CT muscles, then, must very subtly
increase their contraction in order to prevent a rise in intended F0 (singing sharp).
Intensity in the voice source spectra is decreased by decreasing the amplitude of
mucosal oscillation, and that is accomplished by decreasing the amount of
adductory force along with subglottic pressure and aerodynamic flow. With a decrease in subglottic pressure, the
CT muscles must subtly decrease their contraction intensity in order to prevent
a lowering of intended F0 (singing flat). The
adductory-abductory muscles may also decrease their contraction intensity viz a viz the subglottic pressure.
These conditions produce fewer higher
partials in the voice source spectra, compared to optimal adduction in the
upper register coordination. A rather
steep spectral slope results. The F0 is by far the most prominent
partial. Partial amplitude reduces by at
least 12-dB per doubling of the F0, often more. These acoustic characteristics produce
typical contributions to perceived voice quality which may be described as flutiest, lightest, and thinnest when
compared to upper register.
Whistle Register
The
biomechanical details of whistle register coordinations are the least
documented of all the register coordinations (Miller, et al., 2001). The biomechanical coordination of the larynx
that produces flute/falsetto register is a prerequisite for the induction of whistle
register, that is, absence of thyroarytenoid contraction and near maximum
cricothyroid contraction. One visual
observation of whistle register production, based on laryngeal videoendoscopic
images, suggests that an as yet undocumented biomechanical action creates a
suppressed cessation of vocal fold oscillation in the posterior “halves” of the
vocal folds' membranous portions (Personal communication, Robert Bastian, M.D.,
Loyola Voice Institute, Chicago, Illinois, 1999). Only the front “halves” vibrate, therefore,
and produce F0s
that are quite high (in the E6/F6 to C7 range; one octave lower in males) in
what may be described as a tiny voice
quality as opposed to a thinnest voice quality in flute/falsetto.
These conditions
appear to prevent increases and decreases of vibratory amplitude, thus changes
of vocal volume at the respiratory/vocal fold level do not appear to be
possible. Increased conditioning of the
biomechanical coordination and the vocal fold surface tissues and ligaments
might possibly enhance the “full-bodiedness” of this tiny voice quality.
Presumably, perceived volume can be enhanced by vocal tract adjustments,
but they are likely to be minimal due to the necessity for a quite wide
jaw-mouth opening and a very small pharynx.
Current observations are that only some people can produce this
register. Some people with vocal fold
nodules are able to produce it, whereas they are not able to do so with normal
vocal fold tissues (Personal communication, Robert Bastian, M.D.).
Register
Transitions
When people
speak expressively within a
relatively wide range of F0s, most of their F0s will be produced in their lower
register larynx coordinations (thyroarytenoid prominence), but some of their F0s will be produced in the lower F0s
of their upper register coordinations (cricothyroid prominence). That means that they are transitioning
between the two register coordinations.
If there are no audible, abrupt changes in their voice quality when they
transition from one register to the other—only a subtle, blended change—then
that is evidence that their habitual neural networks have enacted a blended or
“melted” transition between the two coordinations. If their voices do produce abrupt changes in
voice quality—often called voice cracks or breaks—then that is evidence that
they do not have habitual neural
networks that enact a blended transition.
The likelihood of voice cracks or breaks is greater when young
adolescents are experiencing voice transformation or when people have inflamed
and swollen or stiffened vocal folds.
|
Figure 7: Graphic description of an abrupt vocal
register transition.
[From
I.R. Titze, Principles of Voice Production. Copyright © 2000, |
|
Transitions between pulse and lower
registers. When transitioning from pulse to lower
register, the cricothyroid muscles engage to assist in the stabilization of the
shorter vocal fold lengths, and thus they participate in the production of
intended F0s. When the cricothyroids are engaged, the
intensity of contraction by the thyroarytenoids increases and that action moves
the vocal fold cover tissues slightly closer to each other. That results in a slight increase of vocal
fold adduction and necessitates a complementary increase in subglottal air
pressure. Aerodynamic flow and mucosal
waving then become more periodic, the temporal gaps in pulse register no longer
occur, and a more sustained vocal sound is perceived. When transitioning from lower to pulse
register, the cricothyroids disengage and subsequent shortening and lengthening
of the vocal folds is carried out exclusively by increases and decreases in the
contraction of the thyroarytenoids (see Titze, 2000; Titze, et al., 1989). When in lower register (shortener prominent),
both of the two primary vibrational modes of the vocal folds are present due to
the thickness and laxness of the vocal fold tissues.
If vocal fry
pulses are perceived as a result of the pulse register coordination, then with
the transition to lower register, mucosal waving converts from pulsed “wave
packets” with temporal gaps to the periodic waving that produces sustained
vocal sound. If the increases of
subglottal pressure, aerodynamic flow, and engagement of the thyroarytenoids
are abrupt, the transition will be perceived as an abrupt crossover from one
quality to another at a particular F0.
If the increases of subglottal pressure, aerodynamic flow, and
engagement of the thyroarytenoids are evenly parceled in very small increments
over several F0s,
the transition will be perceived as a blended change from one quality to
another. If the pulse register
coordination produces perceived sustained tones in singing, then the perceived
transition to lower register coordination is much less obvious; if it is
skilled, the likelihood of audible or even kinesthetic detection is
reduced.
Transitions between lower and upper
registers. In lower register the thyroarytenoid muscles
(shorteners) are more prominently contracted than the cricothyroid muscles
(lengtheners), and the vocal fold cover tissues are in a comparatively shorter,
thicker, and more lax state. When vocally unskilled singers lengthen their
folds to raise the fundamental frequency (ascend in pitch), their lengthener
muscles gradually increase their contraction intensity. If they do not have vocal neural networks
that enable their shortener muscles to complementarily reduce the intensity of
their contracting, then the agonist-antagonist “tug of war” between the two
will become increasingly intense.
Eventually, an impasse will be reached and novice singers will either
stop singing or an abrupt readjustment of contraction intensities between the
shortener and lengthener muscles will occur.
The abrupt adjustment will be to a lengthener prominent state and an
abrupt change in the voice source spectra will occur to produce an audible
voice quality change—to the lighter-thinner
quality that is associated with upper register—along with an abrupt reduction
of vocal volume (as the closer-opener muscles synergistically adapt).
When vocally skilled singers lengthen their
folds to raise the fundamental frequency, their lengthener muscles also gradually
increase their contraction intensity.
But they will have vocal neural networks that enable their shortener
muscles to complementarily reduce the intensity of their contracting. The resulting reduction of bulk in the
thyroarytenoids and the lengthening action of the cricothyroids create a
gradual thinning and tautening of the vocal fold cover tissues. As those changes occur, the vibrational modes
of the vocal fold cover tissues also undergo gradual changes and very subtle
changes of voice source spectra and perceived voice quality occur
correspondingly. The bottom-to-top
waving vibrational mode of the vocal folds gradually diminishes with vocal fold
lengthening and thinning, and voice quality gradually and subtly changes from thicker-more full-bodied toward thinner-lighter even within the overall
shortener prominent state. That means
that within the shortener prominent state
of lower register, there can be a voice quality range that can be described
as thicker-thinner, more and less full-bodiedness, or more and less lightness.
Thus, a subtle,
intricate, agonist-antagonist “give-and-take” between the shortener and
lengthener muscles will enable a gradual crossover transition, over several
pitches, to a lengthener prominent state, and the medial-to-lateral-to-medial-to-lateral
vibrational mode becomes more prominent.
With those changes, the voice source spectra are altered and the lighter-thinner quality that is
associated with upper register is audible.
As lengthening continues, the intermediate and deep layers of the lamina
propria become increasingly stretched and taut and, eventually, only the
superficial layer will be involved in vocal fold tissue oscillations (Titze,
2000) and only the medial-to-lateral-to-medial-to-lateral vibrational mode will
be present. When the closer-opener
muscles produce increased adductory intensity, the top-to-bottom contact area
of the vocal fold cover tissues is increased, and thus voice source spectral
changes occur that produces a perceived voice quality that can be described as
having more full-bodiedness even
within the lengthener prominent state.
That means that within the
lengthener prominent state of upper register, there can be a voice quality
range that can be described as thicker-thinner,
more and less full-bodiedness, or more and less lightness.
When
transitioning from upper register to lower register, a reverse process occurs
in skilled singers, and in approximately the same pitch area.
When
transitioning from upper register to lower register, the unskilled singer is
not likely to have neural networks that will enable a blended transition from
the lengthener prominent state to the shortener prominent state. Either (1) the shorteners will abruptly
engage to create an audible crossover “break” in voice quality with an abrupt
increase in vocal volume, or (2) the lengthener prominent state will continue
into lower and lower pitches. In the
absence, then, of sufficient contraction of the shortener muscles (and their
adductory gesture), the vocal fold margins typically recede away from each
other and a breathy voice quality
becomes audible, and singers’ lowest pitches will not be produced as their
voices “fade out”.
Transitions between upper and
falsetto/flute registers. When transitioning from upper register to
falsetto or flute register, the thyroarytenoid muscles gradually reduce their
contraction to zero so that the cricothyroid muscles assume total influence
over the lengthening and shortening of the vocal folds. The resulting elimination of thyroarytenoid
bulk and the lengthening and thinning action of the cricothyroids create a
thinnest, longest, and most taut range of vocal fold cover tissue states. As F0 increases, the intermediate and deep
layers of the lamina propria become stretched and taut even more extensively
than in the upper register condition.
That means that the only vibrational mode of the vocal folds is the
medial-to-lateral-to-medial-to lateral mode, and the bottom-to-top waving
vibrational mode of the vocal folds may no longer be present. Quite high F0s are possible in this register, as a
result.
In the
falsetto/flute register coordination, the vocal fold cover tissues are so thin
that the top-to-bottom contact area is generally smallest and the oscillating
tissue is in a shallowest depth. The
result is reduced production of harmonics and a proportional predominance of
the F0
among the partials of the voice source spectra (Walker, 1988; Titze,
2000). The perceived voice quality can
be described as thinnest and flutiest.
When
transitioning from falsetto or flute register to upper register, the
thyroarytenoid muscles re-engage but the cricothyroids are predominant. The neuromuscular capabilities of the larynx
(see Endnotes 2 and 3) are such that very subtle engagement of the shorteners
can occur in this register transition.
The increased shortening and the addition of some thyroarytenoid bulk
not only helps to lower the F0, it also reintroduces the spectral
characteristics of the upper register voice source and its contributions to
perceived voice quality.
If the removal
or re-engagement of the thyroarytenoids is abrupt, the register transition will
be perceived as a sudden crossover from one quality to another at a particular
F0. If the changes are evenly parceled in very
small increments over several F0s, the transition will be perceived as a
blended change from one quality to the other, over a region of crossover
frequencies, and the likelihood of audible or kinesthetic detection is
minimized.
Prepubescent
males and females share the same general register characteristics, although
there is wide variety in the coordinations that have been learned. A history of swollen vocal folds from voice
abuse, upper respiratory infections, or other disease states are but a few of
the sources of variation in learned register transitions and pitch range. During pubescent female voice transformation, as the vocal folds lengthen and
thicken, and the trachea increases its dimensions, the basic register
coordinations continue to be present, but neural networks that coordinate
register transitions appear to go through a period of adjustment (Gackle,
2000).
During pubescent
male voice transformation, as the
vocal folds lengthen and thicken, and the trachea increases its dimensions, the
basic upper and lower register coordinations continue to be present IF those
boys have had appropriate pre-adolescent singing experiences. Typically, those experiences instantiate in
vocal neural networks a “template coordination” for blended register transitions
that only require modifications during the laryngeal growth spurts of
adolescent voice transformation. Boys
who have not had appropriate pre-adolescent singing experiences are much more
likely to experience register transition “breaks” and “flip-flops”. Cooksey, et al., (1985; published only in
Cooksey, 2000) produced evidence that male falsetto register first appears in
the high mutation stage of adolescent voice transformation (Midvoice II in the
Cooksey Voice Classification Guidelines).
Transitions between falsetto/flute and
whistle registers. When transitioning from falsetto or flute
register to whistle register, an undocumented biomechanical action is engaged
to suppress mucosal waving in the posterior “halves” of the vocal folds' membranous
portions. As described earlier, only the
front “halves” vibrate and produce F0s that are quite high and even thinner in
quality than flute-falsetto. The
medial-to-lateral-to-medial-to lateral vibrational mode that is present in
falsetto/flute register continues into whistle register. When singers first produce this register, the
transition from flute-falsetto commonly is “unstable” and may be abrupt. With vocal slides from flute-falsetto to
whistle and back again, and with an imagined model of melting the two together,
the transitions may become blended. The
coordinations for sung pitches can then be learned and the two registers can
become melted in those laryngeal coordinations.
Effects
of Vocal Acoustics on Laryngeal Register Adjustments:
“Middle
Register”?
At the time of
the 1982 CoMeT Voice Registers Committee report (previously described), no
measurable evidence for a middle register had been found (Register #2A) and the committee did not support its certain existence. Singing teacher committee members strongly
argued that a middle register exists.
They indicated that middle register (#2A) is thought to result from
“mixtures” of the laryngeal coordinations that are related to chest (#2 or
lower) and head (#3 or upper) registers.
The centuries-old concepts of voce
mista (voix mixte) and the zoni di passaggii were cited as evidence
for the existence of this register.
In 1983, 1984,
and 1988, however, Titze reported studies showing that reverberating subglottal sound waves could influence
reactive, involuntary adjustments in
laryngeal muscle coordinations. Those
adjustments, in turn, resulted in acoustic changes in supraglottic sound
waves. When this phenomenon occurs in
human singers, they and other listeners perceive a sound quality difference and
label it as a register transition. These
register transitions have been recorded in the F0
ranges that are associated
with the zoni di passaggii of the
Italian vocal pedagogy tradition (Miller, 1977). Austin (1992) and Austin and Titze (1997)
have extended those findings, and they appear to explain the middle register in
singing and perhaps other perceived phenomena that are related to vocal
registers, such as “lift points”.
How do reactive
or involuntary register transitions occur?
When vocal sound
is produced, radiating sound pressure waves are created in two opposite
directions. They travel:
1.
upward
through the supraglottic vocal tract and into the surrounding air; and
2.
downward
into the subglottic trachea.
During upright
stance when singing, the trachea is a tube with a relatively stable length of
about 14-cm to 15-cm (Titze, opening presentation, this conference). Its open circumference dimension also is
relatively stable during voicing because of the nearly rigid cartilage rings
that encircle it. Between human beings
of the same age range, tracheal dimensions vary a relatively small amount,
including between males and females.
There is only about a 10% to 20% difference of tracheal length between
the longest in adult males and the shortest in adult females.
Because the
dimension of each person's trachea is relatively stable when standing well, its
resonance frequency also is relatively stable.
According to measurements by Ishizaka, et al. (1976) and Cranen &
Boves (1987), the resonance frequency of average-sized adult tracheas can range
from ± 500-Hz (± C5) to ± 600-Hz (± D5) (see also Titze, 2000). When the vocal folds are closed and
oscillating, there is no opening from which subglottic
sound waves may radiate out and away.
During voicing, then, sound pressure waves in the trachea repeatedly
impact on the underside of the waving
vocal folds.
When the F0 of the waving folds approaches the
resonance frequency of the trachea, the
dimensions of the trachea will effect an increase in the SPL of the
subglottic sound waves. Due to that
intensity gain, the impact of the sound pressure waves on the underside of the vocal folds is
increased. Those repeated impacts
produce interference with vocal fold cover tissue oscillations. That interference has been referred to as acoustic loading or acoustic impedance of vocal fold
oscillations (Rothenberg, 1981a,b; Titze, 1983, 1984, 1988, 2000).
When motor areas within the cerebral cortex have set in
motion the singing of a learned F0 pattern, but one or more of the F0s approach or match the resonance
frequency of the trachea, then the pressure-sensitive mechanoreceptors in the
vocal folds will detect acoustic loading of the continuously oscillating vocal
folds. The interference will be reported
to the brainstem in milliseconds of time.
High-speed involuntary or reflexive motor commands will then be enacted
and sent to the laryngeal muscles to make compensatory coordination adjustments
so that voicing can continue.
In both male and female adult bodies, the involuntary,
reactive adjustments take place in the F0 areas that match the frequencies of the zoni di primo and secondo passaggii, or the frequencies of their prominent
harmonics. The acoustic loading
phenomenon, therefore, can explain the shortener-lengthener muscle adjustments
and the auditory and kinesthetic perception of a middle register. Some laryngeal muscle adjustments can be
subtle and produce a mildly abrupt voice quality change—voluntarily or
involuntarily—that are sometimes referred to as lift points.
Above the vocal
folds, the vocal tract can provide an open end from which vocal sound waves can
radiate. Unlike the trachea, the vocal
tract can vary its dimensions in numerous ways.
Very broadly speaking, two vocal tract cavities can open or narrow: (1)
the pharyngeal cavity (its “throat part”) and (2) the oral cavity (its “mouth
part”).
When the adjustable vocal tract becomes more narrow, then
more and more of the radiating sound wave activity within it will be reflected
onto the topside of the oscillating
vocal fold tissues. Under those
conditions, the vocal folds are receiving increasingly intense pressurized
impacts from both the subglottic and supraglottic sound waves. When this “double-dose” of acoustic loading
occurs, it can be referred to as
acoustic overloading of the vocal
folds (see Thurman & Welch, 2000, p. 446).
During speaking or singing, the general spatial dimensions
of the pharyngeal and oral cavities can be enlarged “too much”, restricted “too
much”, or optimally “opened”.
Inexperienced, unskilled singers will use the only vocal tract
adjustments that they know—the adjustments that are appropriate for
conversational speech. When speaking or
singing in greater-than-speech F0 and intensity ranges, the conversational
speech vocal tract adjustments will result in acoustic overloading by the
subglottal and supraglottal sound pressure waves. Under those conditions, one of two reactive
laryngeal adjustments can take place:
1. over-compensation, that is, increased
contraction intensity in the internal larynx muscles (and typically some of the
external larynx muscles) and increased lung-air pressure, so that the acoustic
interference can be overpowered and continuation of vocal sound can be preserved;
or
2. under-compensation, that is, an abrupt
adjustment of internal laryngeal musculature that produces an abrupt voice
quality change that is referred to as a register “break” or, if the abrupt
adjustment is relatively minimal, a register “lift”.
When experienced, skilled singers sing many pitches and
create many vowel formations of the vocal tract, there is a constant “tuning”
and “re-tuning” between the laryngeal muscle coordinations and the shaping of
the vocal tract in order to maintain sustained vocal sound, vowel
intelligibility, and desired voice qualities (Colton, 1994; Sundberg, 1987;
Titze, 2000).
In both male and
female adult bodies, the involuntary, reactive adjustments of the internal
larynx muscles typically take place in two F0 areas that are approximately one octave
apart. In adult males, they most
frequently can occur between Db3 to F#3, and Db4
to F#4. In adult females,
they most frequently can occur between Db4 to F#4, and Db5
to F#5. These F0 ranges match the frequencies of the
traditional zona di primo passaggio and
the zona di secondo passaggio, or the frequencies of their prominent
harmonics. The acoustic loading
phenomenon, therefore, can explain the auditory and kinesthetic perception of a
lower, a middle, and an upper register.
Some laryngeal muscle adjustments can be more subtle and produce a
mildly abrupt voice quality change—voluntarily or involuntarily—and they could
be what is often referred to as lift
points.
Skilled
adjustments of the pharyngeal and oral cavities, and the internal laryngeal
muscles, occur in singers whose brains have learned
to blend the associated voice quality transitions (Titze, 2000). If, however, the vocal tract's pharyngeal and
oral dimensions are appropriately adjusted during performance of the F0s that approach and match the tracheal
resonance frequency, then tone continuity will be preserved and a subtle,
smoothly blended change of perceived voice quality is much more likely. When these adjustments have been brought into
conscious awareness, they eventually can become learned, habitual, or automatic
motor patterns. A common method of
helping people adjust the “mouth part” of the vocal tract is called vowel modification.
Voluntary, learned neuromuscular coordination changes in the
larynx create the voice source spectra changes that we perceive and refer to as
register transitions. Voluntary
processing is initiated by various areas in the brain's motor cortex. The muscles that are needed to produce chosen
pitches, loudness levels, and voice qualities are recruited and sequenced by
cortical and subcortical motor networks of the nervous system, and they can
induce them both inside and outside conscious awareness.
Human nervous systems can “assemble” automatic, habitual
neural networks (learning) that induce register “breaks”, and they can assemble
automatic networks that induce “melted” or “blended” register transitions, and
induce them in predictable pitch areas. Human nervous systems also are capable
of inducing deliberate register transitions in a variety of pitch areas, and of
changing the habitual transitions to new habitual pitch areas. In other words, voluntary register
transitions can be produced in conscious awareness or outside conscious
awareness, and many register coordination patterns can be learned.
SOME APPLICATIONS OF THIS THEORY OF VOCAL
REGISTERS
TO VOICE EDUCATION AND TO CLINICAL
SETTINGS
Learning How to Sing with Pitch Accuracy
Human auditory processing for spoken and sung
language begins during the third trimester of womb life (Eisenberg, 1969;
Lecanuet, 1996; Panneton, 1985; Pujol, 1993).
During infancy and the early childhood years, human beings hear, see,
and sense language models and their expressive-interactive, imitative, and
exploratory-discovery capability-ability clusters activate to learn spoken and
sung language (DeCasper, et
al., 1994; Meltzoff, 1988a,b; Siegler, 1996).
In
Western cultures, once language is learned, all human beings do nearly all of
their speaking in the lower register (shortener prominent) larynx
coordinations. The lower register family
of voice qualities, therefore, is the most common one in speech. That means that, by far, the most
neuromuscular “practice” in nearly everyone has been the shortener prominent
one. The nervous system, therefore, has
an extensively elaborated array of neural networks to “run” talking in that
coordination, but a fairly undeveloped array of neural networks to “run”
speaking or singing in the lengthener prominent coordination (upper register).
In
most people, therefore, the varied shortener-lengthener stabilizations that are
necessary for reasonable pitch accuracy in singing are most likely to show up
first in the shortener-prominent, lower pitch range area. But when people attempt to sing gradually
higher pitches starting from there, they may not have an array of neural
networks that would enable them to transition “smoothly” from shortener
prominent to lengthener prominent coordinations (upper register). A first pitch area to begin learning pitch
accuracy in would be, therefore, in lower register, using simple limited
pitch-range songs (for pre-adolescent children and adult females: a pitch range compass of about a perfect 5th
or a 6th, e.g., “Go Tell Aunt Rhody” or the chorus of “Jingle Bells”
in the key of Ab, with the melody beginning on C4).
While
having lower register singing experiences, less skilled singers can be led
through vocal sound-making and language-making that involve lengthener
prominent coordinations. For example,
vocal pitch slides and glides (a “yooooo hoooooooooooo” call, modeled by an
adult)and siren sounds and short word phrases can be experienced in a game or
story setting or “just for the fun of it”.
Doing those vocal sound-makings over time result in development of some
foundational neural networks that eventually can be used for singing pitch
patterns in upper register. After
experiencing the “yoo hoo” call enough times, that way of making sounds can be
referred to as the “yoo hoo part of your voice”.
When
relatively accurate pitches are sung in lower register coordinations, and some
experience has been gained with upper register coordinations, then a teacher
might say, “I wonder what would happen if we sang this song in the “yoo hoo”
part of our voices?” There is a high
probability that learners then will be able to accurately sing the same song in
the same key, but pitched one octave higher (melody beginning on C5
in upper register).
If
those songs are sung in the key of C with the melody beginning on E4,
however, inexperienced singers, typically, will sing inaccurate pitches or in
another key right away. That pitch area
is in the transition area between the shortener prominent and the lengthener
prominent neuromuscular coordinations.
The clearly shortener prominent and clearly lengthener prominent
coordinations are relatively gross motor skills. Transitioning between the two requires neural
networks that can deal with a large array of subtle intensities of neuromuscular
contraction in the vocal fold shortener and lengthener muscles, and
inexperienced singers are not likely to have developed those fine motor skill
neural networks yet.
A
way to help inexperienced singers begin to master the register transition
skill: After singing the song in the
upper octave key (upper register), repeat the song in keys that descend from
there in whole or half steps until they pass through the transition area
(usually between F4 and D4). Starting out in the lengthener prominent
coordination optimizes the chances that the vocal fold shortener muscles will
not “overpower” the lengthener muscles and take the pitch “off target”. Their chances of singing with reasonable
pitch accuracy through the transition area will be optimized as a result. If inexperienced singers start in the
lower-octave key and the keys are gradually raised by half or whole steps, they
most likely will continue to engage the shortener prominent register
coordinations into the higher keys, and will start singing below the accurate
pitches, and eventually will go “off tune”.
“Belt” Singing
Belting or belt
quality is a term that was coined in the American musical theatre and was
popularized by the singing of Ethel Merman in the 1940s and 1950s. That style of singing is a staple of musical
theatre in Western civilization. But for
thousands of years, children, adolescents, and adults of nearly all the world's
cultures have sung their folk and popular musics in a strong “belted way”. Current popular and religious musical styles
that have roots in the African-American experience preponderantly use belted
singing (spirituals, blues, jazz, gospel, rock, and so forth).
With the trend toward multicultural music education and
multicultural choral singing comes the necessity for stylistic
authenticity. The vocal qualities that
sounded when “folk” created a culture's sung music are integral to its
expressive style. Change the vocal
qualities and you change the very core of its human expressiveness. It is no longer that culture's music.
Any form of
strong, high intensity (loud) singing involves quite strenuous laryngeal muscle
use and high impact and shear stresses on the vocal folds. So-called belted singing and Western opera
style singing are two forms of strong, high intensity singing (Estill, 1988). There are inefficient, overly strenuous and
fatiguing ways to produce strong singing, including belt quality, and there are
efficient, optimally vigorous ways.
Based on this
paper’s theory of vocal registers, strong belted singing involves maintaining variations
of a shortener and closer prominent coordination of internal larynx muscles
(with higher lung-air pressures) into higher and higher pitches. Thus, the “tug-of-war” tension between the
shortener and lengthener and the closer-opener muscles becomes increasingly
intense as pitches rise. Subtle,
intricate variations in those laryngeal muscle adjustments can produce a
variety of subtle “thickness-thinness” qualities. In order to avoid acoustic overloading of the
vocal folds, the mouth part of the vocal tract must gradually widen as pitches
rise, becoming quite wide open even in the middle of singers’ capable pitch
range. In addition, subtle, intricate
variations in vocal tract adjustments can produce a variety of subtle
“fuller-brighter” qualities. Lifetime
vocal health is possible when:
1. all fundamental vocal skills have been
learned with optimum efficiency (especially including development and
conditioning of the upper and falsetto/flute register coordinations);
2. the
laryngeal muscles, connective tissues, and vocal fold cover tissues are well
conditioned;
3. singers
know how to protect their voices (recovery time, hydration, and the like; see
below).
Applications in Clinical Voice Therapy
Register
neuromuscular coordination tasks (examples described below) can be used to
differentially distinguish between: (1)
the presence of a vocal pathology, (2) deconditioning of the larynx muscles
(thyroarytenoids, in particular), and (3) underdevelopment of vocal
abilities.
Presence of vocal pathologies.
Register neuromuscular coordination tasks can be used to assess the functional viability of the
superior and recurrent laryngeal motor nerves. These assessments relate to conditions such
as laryngeal paresis or paralysis and deconditioning. Register coordination tasks also can be used to assess the severity of inflammatory
swelling of vocal fold cover tissues and other organic pathological disorders
of the vocal folds (e.g., nodules, polyp, cyst, hemorrhage).
In the healthy,
well conditioned larynges of skilled singers, the absence of finely coordinated
register transitions may indicate early neurological disease. For example, a well trained, well conditioned
singer was seen at
Neuromuscular
instabilities were observed visually and aurally during her register
transitions, and that was the only abnormality that was observed during her
voice evaluation. This condition
persisted despite several weeks of voice therapy and could not be explained by
any voice function diagnosed. She was
referred for neurological examination, and was diagnoses with early stages of
multiple sclerosis.
Register neuromuscular
coordination tasks that progressively lengthen and thin the vocal fold cover
tissues (upper, falsetto, and flute register coordinations) can reveal the
degrees of severity in vocal fold cover tissue changes such as swelling and/or
the presence of organic lesions (e.g., nodules, polyp, cyst, hemorrhage,
sulcus). When these conditions are
present, the following common consequences are likely:
1. generally
increased air leakage (breathy quality)
2. voice onset
delays and aphonic episodes
3. generally
increased laryngeal and respiratory effort
4. abrupt or
unstable “flip-flop” voice quality characteristics at register transitions
Among singers
and actors, common contributors to the development of swelling and other
pathological voice disorders is inappropriate use of voice register
coordinations that produce high impact and shear stresses on the vocal fold
mucosa. Using such coordinations with a
larynx that is underconditioned for such higher levels of “stress-demand”, can
also contribute to these disorders.
Deconditioning/underconditioning.
When extensive and/or vigorous voice use has been reduced over time, or
rarely undertaken, atrophic processes occur in the laryngeal nerves and
muscles, and a “softening” occurs in vocal fold tissues (described in Plot
Setting 6). Reduction in the thickness
of the thyroarytenoid muscles (shorteners) results in a recession of the vocal
fold cover tissues away from a complete adduction at the midline. Laryngeal videoendoscopy reveals a gap
between the vocal folds during softer voicing and irregularities in mucosal
waving patterns. These tissue changes
result in reduction of vocal abilities, including register transition
instabilities (e.g., “flip-flops”), as well as decrements in vocal pitch
accuracy, vocal volume, and voice quality.
These decrements in vocal ability are not pathological, per se, but can lead experienced
vocalists to assume that they have a disordered voice that requires vocal
rest. In the case of otherwise healthy
larynx muscle and vocal fold tissues, gradual increases in the extent and vigor
of voice use is needed for a return to normal voicing, along with attention to
blended register transition coordinations.
Underdeveloped vocal abilities.
When a patient’s history does not include voice education experiences,
or includes relatively minimal voice education, then vocal register
coordinations are likely to be “all or nothing”. For example, the lower register coordinations
that are used to produce conversational speech will be strongest and also will
be more likely to be used to produce higher-range pitches, especially pitches
that have greater vocal volume. When
voice is used that way frequently, the risk of vocal fold swelling and other disorders becomes
greater. Also in less skilled vocalists,
the fine motor skills that are required to produce blended or smooth register
transitions are less likely to be present.
Correlating a complete vocal health history, diagnostic vocal tasks, and
laryngeal videoendoscopy can be used to differentially distinguish between
underdeveloped vocal abilities and deconditioning, and vocal pathology
conditions.
Basic register neuromuscular coordination
tasks: Patients can be asked to imitate vocal
sounds or word phrases as modeled by clinicians in their lower register (e.g., “Whoooooo are youuuuuu?” or “say the
word ‘Hello’ in three different and interesting ways”). The phrases would be presented in two or
three prosodic variations that necessitate changes of pitch, volume, timing,
and voice quality. This task especially
evaluates basic neuromuscular viability of the recurrent laryngeal motor nerves
that innervate the vocal fold opener-closer-shortener functions (shorteners
more prominently contracted than lengtheners).
Then patients
can be asked to imitate the clinician’s upper
register word phrases (e.g., the call, “Yoo hoo”) in two or three prosodic
variations. Patients can then be asked,
“How close can you come to starting in the
‘yoo-hoo’ part of your voice and slide downward continuously into the
‘who-are-you’ part of your voice?” Then:
“How close can you come to starting in the ‘who-are-you’ part of your voice and
slide upward continuously into the ‘yoo-hoo’ part of your voice?” This task
especially evaluates basic neuromuscular viability of the superior laryngeal
motor nerves that innervate the vocal fold lengthening functions in upper
register (lengtheners more prominently contracted than shorteners).
Finally,
patients can be asked to imitate the sound of a newly born, tiny puppy, as
modeled by the clinician, and afterward, to create sound spirals that ascend
“wherever they might want to go”.
Puppy-cry sounds are intended to elicit falsetto register in males and
flute register in females. This task especially evaluates basic neuromuscular
viability of the superior laryngeal motor nerves that innervate the vocal fold
lengthening functions in falsetto and flute registers (lengtheners only).
Patients who
sing, can be asked to sing 5-4-3-2-1 musical scale pitch patterns that begin in
lower register and descend to near the lowest producible pitch. The beginning pitch can be about F3
for males and F4 for females.
Each succeeding scale pattern can begin on a pitch that is a semitone or
whole tone below the previous starting pitch.
Singers can then
be asked to sing 5-4-3-2-1 musical scale pitch patterns that begin in upper
register and descend until lower register is clearly sounding. The beginning pitch can be about C4
for males and D5 for females.
Each succeeding scale pattern can begin on a pitch that is a semitone
below the previous starting pitch.
Singers can then
be asked to sing 5-4-3-2-1 musical scale pitch patterns that begin in upper
register and ascend to their highest producible pitch. The beginning pitch can be about C4
for males and D5 for females.
Each succeeding scale pattern can begin on a pitch that is a semitone above the previous starting pitch.
Bastian, Keidar,
and Verdolini-Marston (1990) validated two pitch pattern tasks that can be
repeated several times in progressively higher keys (vocal fold thinning) to
evaluate the severity of vocal fold swelling.
One of the patterns is the first phrase only of “Happy Birthday” and it
must be sung very softly and thinly (pianissimo
or pp; and tiny-sounding, thus adding
a greater degree of vocal fold thinness).
Changed-voice males must sing in falsetto register and the first pitch
would be C4, and the key of each repetition would be higher by one
semitone, until the highest pitch is C5. Females and unchanged males would begin on C5
in upper register and the key of each repetition would be higher by one
semitone, until the highest pitch is C6. Clinicians and patients can use this task to
track the progress of swollen vocal folds toward normal dimensions, including
post-surgical healing. For patients who
are uncomfortable with singing, the puppy-cry spirals described earlier can be
substituted.
ENDNOTES
1 A wide variety of neuromuscular coordinations occur to produce all vocal
phenomena. Are speaking and singing so
categorically different, really? To
accomplish both of the so-called speaking
voice and singing voice, human
beings must inhale air into the lungs, close the vocal folds, “squeeze” on the
lungs to pressurized the air therein to create a breathflow that causes the
vocal folds to oscillate, configure the vocal folds and “shape” the vocal tract
in various ways to create a variety of verbal and nonverbal expressions. The only differences appear to be: (1) In speech, vocal fold length is nearly
always in variable flux patterns that produce “sliding” vocal fundamental
frequencies. In singing, a variety of
stabilized vocal fold lengths produce consecutive sustained fundamental
frequencies. (2) Most of the time in
singing, a greater range of fundamental frequencies (pitches), sound pressure
levels (volumes), and radiating spectra (voice qualities) are produced,
compared to speech.
Presumably, this
categorical distinction is instantiated within somewhat differentiated neural
networks that one highly respected neuroscientist refers to as cognits (Fuster, 2003). Among English speaking people, the speaking voice and singing voice cognits are sometimes so differentiated that they
have unfortunate consequences for human beings.
Examples: (1) Some music teachers wonder why their
diagnosed nodules have affected their speaking
voices (hoarseness) but not their singing
voices. (2) A choir sings a
selection that includes both singing and speaking sections. Their singing sounds vocally efficient and is
expressive, but when the somewhat loud speaking section is performed, the
predominant voice quality is pressed, edgy, and harsh. The conductor explains that the singers use
their “natural” speaking voices for
that section, and reveals that he only knows how to teach the singing voice, not the speaking voice. (3) Some otolaryngologists have used the
diagnostic term singer’s nodules,
even though the way singers use their voices for speech may be the most
significant factor in the formation of nodules.
(4) People who are not familiar with the jargon of the voice professions
have arrived for appointments with voice
teachers, expecting to improve tgheir voices for greater effectiveness as
business executives or salespersons.
They were puzzled and dismayed when they were told that the teacher did
not teach the speaking voice, only
the singing voice.
This type of
distinction is not used in any other area of human neuromuscular activity. We do not say that we have walking legs and
running legs, or pushing arms and pulling arms.
Do we have cognitive brains, affective brains, and sensorimotor
brains? More particularly, do we have speaking registers and singing registers that are categorically
different from each other and require categorically distinct nomenclature?
2 Reflexive motor functions are initiated
by very high-speed sensory input and the circuits are short. Peripheral sensory reception is
delivered: (1) to the spinal column
which immediately triggers spinal motor nerves with no other central nervous
system processing, or (2) to brainstem nuclei which immediately trigger cranial
motor nerves with no other central nervous system processing.
Both spinal and cranial motor neurons extend their
myelinated, largest-diameter axons outward to their target muscles and form
their part of the peripheral nervous system.
At the surface of a target muscle, each axon divides into multiple
terminal branches. Each single terminal
branch is attached to one of the many
muscle fibers that make up that whole muscle.
The multiple terminal branches that extend from one axon, and all of the
muscle fibers that they innervate, are referred to as a motor unit (Burke, 1981;
Vander, et al., 1994, pp. 315-317). The
anatomical point at which an axon terminal branch and a muscle fiber interface
is referred to as a neuromuscular
junction. The muscle fibers that are
innervated by the terminal branches of one neuron are not located adjacent to
each other, but are distributed throughout the target muscle. When a single motor neuron “fires”, all of
the muscle fibers to which it is attached will contract, so that the number of
motor units that are activated relates to the contractile properties of the
whole muscle (more later). The four
motor unit types are described in the table.
Each muscle also has the two types of sensory receptors (affectors).
One type detects degrees of
contraction intensity (Golgi tendon organs), and the other detects degrees of stretch or lengthening
(muscle spindle stretch receptors). The
peripheral end of each sensory neuron's axon is divided into multiple terminals
that receive stimulation from muscle or ligament fibers. Each sensory neuron's cell body is located
astride its axon, and its other end is connected to the spinal or brainstem parts
of the CNS.
There are three types of muscle contractile properties: (1) degree of generated force (related to muscle strength), (2)
degree of contractile speed (related
to quickness of response), and (3) degree of endurance (related to central nervous system or peripheral
neuromuscular fatigability). The degree
of force that is generated by a
single motor unit depends on (1) the size of the neuron and (2) the number of
impulses (action potentials) that are generated per second. Usually, larger neurons generate more force
and smaller neurons generate less force.
More impulses per second generate more force.
When a motor unit has higher numbers of terminal branches,
and therefore innervates more muscle fibers, it is regarded as a large motor
unit. Motor units with small numbers of
terminal branches and innervated muscle fibers are regarded as small motor
units. Muscles that are involved in
relatively coarse motor coordinations (back and legs, for example) are
generally large and have relatively few motor units, and each motor unit may
control hundreds to even thousands of muscle fibers. On the other hand, muscles that are capable
of participating in intricate, fine, subtle, or delicate motor coordinations
(hands, eyes, and larynx, for example) tend to be small, and they have numerous
motor units that may control only one to a few muscle fibers.
The central nervous system can very gradually increase the
contractile force of small muscles, that have many motor units, by increasing
the number of activated motor units in very small increments. This motor unit activation process is called recruitment of motor units. Another means by which the CNS can alter the
contractile forces of muscles is by increasing or decreasing the frequency with
which action potentials course
through a muscle's motor units. These
variations of motor unit recruitment and action-potential frequency are capable
of producing highly intricate coordination patterns in multiple
agonist-antagonist muscle pairs.
When muscles are contracted toward their maximum intensity,
the CNS recruits motor units in a specific order (Gordon & Patullo, 1993;
Williams, et al., 1987). The slow and
fatigue resistant motor unit types (S) are recruited first, followed by the
fast and fatigue resistant types (FR), then the fast and fatigue intermediate
types (FInt), and finally the fast and fatigable types (FF). When muscles are reducing the intensity of
their contracting, the CNS reduces the number of activated motor units by
deactivating them in the reverse order from FF to FInt, to FR, to S. These processes can be accomplished in very
small increments within smaller muscles that have a high ratio of small motor
units in them. As higher intensity and
finer-tuned use is repeated, the metabolic processing of fast-fatigable
glycolytic motor units is converted more and more to fast and fatigue resistant
oxidative processing.
Initially, physiologists categorized skeletal muscle fibers
into two types, according to their speed
of contraction and the extent of their resistance
to fatigue (see Table 2; Vander, et al., 1994, p. 327). Slow-twitch
muscle fibers (Type I) contract at slow speeds and also are resistant to
fatigue. They can continue to contract
for long periods of time and, therefore, require a rich capillary blood supply
to deliver their primary fuel, oxygen.
They have been referred to as red
muscle fibers (the dark meat in chicken, for instance). Fast-twitch
muscle fibers (Type II) contract at high speeds and do not use oxygen as
their primary fuel. They have a
comparatively minimal blood supply and have been referred to as white muscle fibers. Their primary energy source is glucose, used
in the form of glycogen. Glycogen is
stored in the body through metabolic processes and can be depleted more rapidly
(O2 is delivered by the respirocardiovascular
system and is continually renewable).
White, glycolytic muscle fibers, therefore, fatigue faster. More recently, two versions of Type II muscle
fiber were labeled as Types IIa and IIb (see table). Nearly all muscles, including the internal
and external laryngeal muscles, contain all fiber types and all four related
motor unit types (Vander, et al., 1994, p. 327).
3
During learning, the organizing phase
of vocal motor functions occurs in various areas of the frontal lobes such as
Broca's area in the premotor cortex, and supplementary motor areas, which are
inter-looped with the basal ganglia, cerebellum, thalamus, the limbic system,
and the sensory networks. The execution phase of those functions
begins in the vocalization areas of the two primary motor cortices and extends
downward through axons that form the corona radiata, and eventually through the
periaqueductal gray (PAG) area of the midbrain and brainstem to the nucleus
ambiguus (located within the medulla oblongata), and finally through the right
and left vagus nerves to the relevant muscles (see Figure 3 and Holstege,
1996). Thus, the reflexive vocal neural
networks of the nucleus ambiguus are entrained by higher brain areas to
complete the enactment of learned vocal coordination patterns. When learned functions have been repeated a
sufficient number of times, frontal motor areas decrease their activation as
the subcortical areas become more elaborated and sensitized.
When the two vagus nerves extend from the brainstem, they
are actually made up of a few hundred thousand long axons that are sheathed
together. The axons of laryngeal motor neurons (effectors)
extend outward from their cell bodies in the medulla's nucleus ambiguus. They eventually branch off from the main vagus
nerve trunk and extend to their target muscles (see Figure 3). Each muscle of the larynx also has the two
types of sensory receptor fibers (Golgi tendon organs and muscle spindles). Laryngeal
sensory neurons extend from the target muscles, join the vagus nerves that
extend into the brainstem where they synapse with central nervous system
sensory neurons in the nucleus ambiguus.
From there, sensory signaling is distributed to a variety of processing
areas within the brain.
4
Sensory reception networks for one’s own voice receive feedback about a
“running” series of learned vocal coordinations, and report that feedback to
“interested” brain areas for “interpretation.”
For example, “status reports” are sent to various brain areas about the
stretching force exerted on muscles, degrees of subglottic pressure, and
relative positioning of the laryngeal cartilages (Larson, 1988; Wyke,
1983b). This feedback may be used to
adjust elements of ongoing coordination sequences, or to change subsequent
sequences so that they may more closely approximate a target intention. Auditory
feedback and kinesthetic feedback
participate significantly in the motor adjustments of trained singers and
speakers, but much less so in less-trained singers or speakers (see, e.g., Thurman
& Welch, 2000, Book I, Chapter 6 for a brief review, and Ward & Burns,
1978). Most auditory and nearly all
kinesthetic feedback is processed outside conscious awareness (implicit
perception, see Thurman & Welch, 2000, Book I, Chapter 7 for a brief review).
High-speed laryngeal capability is entrained and refined by
singers who learn how to sing very rapid and wider-interval pitch patterns--the
melismas of Baroque, jazz, and African-American gospel musics, for
instance. The true extent of that capability
is realized only when nearby unnecessary muscles are released from interfering
contraction. When unnecessary muscles interfere, they force a slowing of
higher-speed laryngeal muscle movement.
Capabilities for laryngeal muscle speed and fatigue resistance
change with:
1. morphological
age (compare early childhood and adolescent voice transformation
capabilities with mature adult capabilities);
2. extent
and manner of use (compare vocally healthy and well trained professional
speakers and singers with untrained, quiet conversationalists); and
3. neuromuscular
impairment (neuromuscular disease or injury).
REFERENCES
AND SELECTED BIBLIOGRAPHY
Alipour, F., & Scherer, R.C. (2000). Vocal fold bulging effects on phonation using
a biophysical computer model. Journal of Voice, 14(4), 470-483.
Allen, J.H.
(1935). The Technique of Modern Singing.
Appelman, R.
(1967). The Science of Vocal Pedagogy.
Ardran, G.M., & Wulstan, D. (1967). The
alto or countertenor voice. Music and Letters,
48(1), 21.
Bakken,
R.J. (1991). An overview of laryngeal
function for voice production. In R.T.
Sataloff (Ed.), Professional Voice: The
Science and Art of Clinical Care (pp. 19-47).
Basterra,
J., Dilly, P.N., & Martorell, M.A. (1989).
The autonomic innervation of the human vocal cord: Neuropeptides. Laryngoscope,
99, 293-296.
Bastian, R.W.,
Keidar, A., & Verdolini-Marston, K. (1990).
Simple vocal tasks for detecting vocal fold swelling. Journal
of Voice, 4(2), 172-183.
Bastian, R.W.,
Klitzke, C., & Thurman, L. (2000).
Vocal fold and laryngeal surgery.
In L. Thurman & G. Welch, G. (Eds.),
Bodymind and Voice: Foundations of
Voice Education (Rev. Ed., pp. 620-631).
Bendiksen,
F.S., Dahl, H.A., & Teig, E. (1981).
Innervation types of muscle fibers in the human thyroarytenoid
muscle. Acta Otolaryngology (
Blomgren,
M., & Chen, Y. (1998). Acoustic,
aerodynamic, physiologic and perceptual properties of modal and vocal fry
registers. Journal of the Acoustical Society of
Broad, D.J. (1973). Phonation.
In F.D. Minifee, T.J. Hixon & F. Williams (Eds.). Normal
Aspects of Speech, Hearing and Language (pp. 127-167).
Browne, L.,
& Behnke, E. (1884). Voice, Song, Speech.
Burke, R.E. (1981). Motor units: Anatomy, physiology, and
functional organization. In J.M.
Brookhart & V.B. Mountcastle (Section Eds.),
Castellengo, M.,
Roubeau, B., & Valette, C. (1985).
Study on the acoustical phenomena characteristic of the transition
between ++++++. In
Askenfelt Proceedings of the
Claassen, H.,
& Werner, J.A. (1992). Fiber
differentiation of the human laryngeal muscles using the inhibition
reactivation myofibrillar ATPase technique.
Anatomy and Embryology, 186,
341-346.
Cooksey, J.
(200). Voice transformation in male
adolescents. In L. Thurman & G.
Welch (Eds.), Bodymind and Voice:
Foundations of Voice Education (Rev. Ed., pp. 718-738).
Cooksey, J.M.,
Beckett, R.L., & Wiseman, R. (1985).
A longitudinal investigation of selected vocal, physiological, and
acoustical factors associated with voice maturation in the junior high school
male adolescent. Unpublished research
report,
Cooper, D.S.,
Partridge, L.D., & Alipour-Haghighi, F. (1993). Muscle energetics, vocal efficiency, and
laryngeal biomechanics. In I.R Titze
(Ed.), Vocal Fold Physiology: Frontiers
in Basic Science (pp. 37-92).
Cranen, B.,
& Boves, L. (1987). On subglottal
formant analysis. Journal of the Acoustical Society of
DeCasper, A.J.,
Lecanuet, J.-P., Busnel, M.-C., Granier-Deferre, C., & Maugeais, R.
(1994). Fetal reaction to recurrent
maternal speech. Infant Behavior and Development, 17, 159-164.
Duey, P.
(1951). Bel Canto in Its Golden Age.
Elder,
G.C.B., Bradbury, D., & Roberts, R. (1982).
Variability of fiber type distributions within human muscles. Journal
of Applied Physiology, 53(6), 1473-1480.
Emile-Behnke, K.
(1945). The Technique of Singing.
Estill, J.
(1982). The control of voice
quality. In V.L. Lawrence
(Ed.). Transcripts of the Eleventh Symposium: Care of the
Professional Voice (pp.
152-168).
Estill, J.
(1988). Belting and classic voice
quality: Some physiological differences, Medical
Problems of Performing Artists, 3, 37-43.
Estill, J., Baer, T., Harris, K.S. & Honda, K.
(1985). Supralaryngeal activity in a study of six voice qualities. In A. Askenfelt, S. Felicetti, E. Jansson,
& J. Sundberg (Eds.), Proceedings of
the Stockholm Music Acoustics Conference (pp. 157-174).
Estill, J.,
Baer, T., Honda, K. & Harris, K.S. (1983).
The control of pitch and voice quality:
An EMG study of supralaryngeal muscles,
In V.L. Lawrence (Ed.). Transcripts
of the Twelfth Symposium: Care of the Professional Voice (pp. 86-91).
Estill, J.,
Baer, T., Honda, K. & Harris, K.S. (1984).
The control of pitch and voice quality:
An EMG study of infrahyoid muscles,
In V.L. Lawrence (Ed.). Transcripts
of the Thirteenth Symposium: Care of the Professional Voice (pp.
65-69).
Fröschels, E.
(1920). Singen und Sprechen. Leipzig:
Deuticke.
Fuster, J.M. (1997).
The Prefrontal Cortex: Anatomy, Physiology, and
Neuropsychology of the Frontal Lobe
(3rd Ed.).
Fuster, J.M.
(2003). Cortex and Mind: Unifying Cognition.
Gackle, L.
(2000). Understanding voice
transformation in female adolescents. In
L. Thurman & G. Welch (Eds.), Bodymind
and Voice: Foundations of Voice Education (Rev. Ed., pp. 739-744).
Garcia, M.
(1854-1855). Observations on the human
voice.
Proceedings of the Royal Society of
Gordon, T., & Pattullo, M.C. (1993). Plasticity
of muscle fiber and motor unit types. Exercise and Sports Sciences Reviews,
21, 331-362.
Graney, D.O.
& Flint, P.W. (1993). Anatomy. In C.W. Cummings & J.M. Fredrickson
(Eds.), Otolaryngology--Head and Neck
Surgery, Vol. 3: Larynx/Hypopharynx (2nd Ed., pp. 1693-1703).
Garcia, M.
(1984). Hints on Singing.
Garrett, J.D., & Carson, C.R. (1991). Neurology
of the laryngeal system. In C.N. Ford
& D.M. Bless (Eds.), Phonosurgery:
Assessment and Surgical Management of Voice Disorders.
Herzel, H.,
& Reuter, R. (1997). Whistle
register and biphonation in a child’s voice. Folia Phoniatrica et Logopedica, 49(5),
216-224.
Hess, M.M.,
& Ludwigs, M. (2000).
Strobophotoglottographic transillumination as a method for the analysis
of vocal fold vibration patterns. Journal of Voice, 14(2), 255-271.
Hirano, M.
(1988). Vocal mechanisms in singing:
Laryngological and phoniatric aspects. Journal of Voice, 2(1), 51-69.
Hirano, M.,
Ohala, J., & Vennard, W. (1969). The
function of the laryngeal muscles in regulating fundamental frequency and
intensity of phonation. Journal of Speech and Hearing Research,
12, 616-628.
Hirano, M.,
Vennard, W., & Ohala, J. (1970).
Regulation of register, pitch, and intensity of voice: An
electromyographic investigation of intrinsic laryngeal muscles. Folia
Phoniatrica, 22, 1-20.
Hollien, H.
(1974). On vocal registers. Journal
of Phonetics, 2,125-143.
Hollien, H.
(1985). Report on vocal registers. Proceedings of the
Hollien, H., & Michel, J.F. (1968). Vocal
fry as a phonational register. Journal of Speech and Hearing Research, 11,
600-604.
Hollien, H.,
& Schoenhard, C. (1983a). The riddle
of the middle register. In
Hollien, H.,
& Schoenhard, C. (1983b). A review
of vocal registers. In V.L. Lawrence
(Ed.). Transcripts of the Twelfth Symposium:
Care of the Professional Voice (pp. 1-6).
Holstege, G.,
Bandler, R. & Saper, C.B. (Eds.) (1996).
Progress in Brain Research, No.
107: The Emotional Motor System.
Howard, D.M.
(1995). Variation of
electrolaryngographically derived closed quotient for trained and untrained
adult female singers. Journal of Voice, 9(2), 163-172.
Howard, D.M.,
Lindsey, G.A., & Allen, B. (1990).
Towards the quantification of vocal efficiency. Journal
of Voice, 4(3), 205-212 [See also Errata, (1991), Journal of Voice, 5(1), 93-95.]
Howard, D.M.,
& Rossiter, D. (1992). Results from
a pilot longitudinal study of
electrolaryngo-graphically
derived closed quotient for adult male singers in training. Proceedings
of the
Howard, D.M.,
Welch, G.F., & Penrose, T. (2001).
Case study acoustic and voice source evidence for the existence of
sub-registers in the countertenor voice.
In T. Murao, Y. Minami, & M. Shinzanoh (Eds.), Proceedings of the 3rd Asia-Pacific Symposium on Music
Education Research and International Symposium on ‘Uragoe’ and Gender (pp.
127-131).
Huttenlocher,
P.R. (1994). Synaptogenesis in human
cerebral cortex. In G. Dawson, &
K.W. Fischer (Eds.), Human Behavior and
the Developing Brain (pp. 137-152).
Ishizaka, K.,
Matsudaira, M., & Kaneko, T. (1976).
Input acoustic impedance measurement of the subglottal system. Journal
of the Acoustical Society of
Keidar, A.
(1986). Vocal register change: An
investigation of perceptual and acoustic isomorphism. Unpublished Ph.D. dissertation,
Keidar, A.,
Hurtig, R., & Titze,
Koike, Y.
(1981). Sub- and supra-glottal pressure
variation during phonation. In K.N.
Stevens & M. Hirano, (Eds.), Vocal
Fold Physiology (pp. 181-191).
Large, J.W.
(Ed.) (1973). Vocal Registers in Singing.
Larson, C.
(1988). Brain mechanisms involved in the
control of vocalization. Journal of Voice, 2(4), 301-311.
Lecanuet, J.-P.
(1996). Prenatal auditory
experience. In
Lecanuet, J.-P.,
Granier-Deferre, C., & Busnel, M.-C. (1989). Differential fetal auditory reactiveness as a
function of stimulus characteristics and state.
Seminars in Perinatology, 13,
421-429.
Lecanuet, J.-P.,
Krasnegor, N., Fifer, W.P., & Smotherman, W.P. (Eds.) (1995). Fetal
Development: A Psychobiological Perspective.
Lindestad, P.,
& Södersten, M. (1988). Laryngeal
and pharyngeal behavior in countertenor and baritone singing--a
videofiberscopic study. Journal of Voice, 2(2), 132-139.
Lindestad, P.,
Fritzell, B., & Persson, A. (1990).
Evaluation of laryngeal muscle function by quantitative analysis of the
EMG interference pattern. Acta Otolaryngologica (
Mackworth-Young,
G. (1953). What Happens in Singing.
Mårtensson, A.,
& Skoglund, C.R. (1964). Contraction
properties of intrinsic laryngeal muscles.
Acta Physiologica Scandinavia,
60, 318-336.
Martin, F.,
Thumfart, W.F., Jolk, A., & Klingholz, F. (1990). The electromyographic activity of the
posterior cricoarytenoid muscle during singing.
Journal of Voice, 4, 25-29.
McAlister, A.
(1997). Acoustic, Perceptual and Physiological Studies of Ten-Year-Old
Children's Voices.
Meltzoff, A.N.
(1988a). Infant imitation and memory:
Nine-month-olds in immediate and deferred tests. Child
Development, 59, 217-225.
Meltzoff, A.N.
(1988b). Infant imitation after a
one-week delay: Long-term memory for novel acts and multiple stimuli. Developmental
Psychology, 24, 470-476.
Merkel, C.
(1863). Anatomie und Physiologie des menschlichen Stimm- und Sprachorgans.
Miller, D.G.,
Schutte, H.K., & Hess, M.M. (2001).
Physical definition of the “flageolet register”. Journal
of Voice, 7(3), 206-212.
Miller, R.
(1977). English, French, German and Italian Techniques of Singing.
Miller, R.
(1986). The Structure of Singing: System and Art in Vocal Technique.
Mori, R.M.
(1970). Coscienza della Voce nella Scuola Italiana di Canto (Awareness of
Voice in the
Mörner, M.,
Fransesson, N., & Fant, G. (1964).
Voice register terminology and standard pitch. Speech
Transmission Laboratory Quarterly Status Progress Report, 4, 12-15.
Murry, T., Xu,
J.J., & Woodson, G.E. (1998).
Glottal configuration associated with fundamental frequency and vocal
register. Journal of Voice, 12(1), 44-49.
Orlikoff, R.F.
(1991). Assessment of the dynamics of
vocal fold contact from the electroglottogram.
Journal of Speech and Hearing
Research, 34, 1066-1072.
Panneton, R.K.
(1985). Prenatal Auditory Experiences
with Melodies: Effects on Postnatal Auditory Preferences in Human
Newborns. Unpublished Ph.D.
dissertation,
Pujol, R. (1993).
Développement et plasticité du système auditif de l'enfant (Development
and plasticity in the auditory system of the infant). Communiquer, 111, 13-16.
Pujol, R., &
Uziel, A. (1986). Auditory development:
Peripheral aspects. In P.F. Timeras
& E. Meisami (Eds.), Handbook of
Human Biological Development.
Richards, D.S.,
Frentzen, B., Gerhardt, K.J.,
Rothenberg, M.
(1981a). Acoustic interaction between
the glottal source and the vocal tract.
In K.N. Stevens & M. Hirano, (Eds.), Vocal Fold Physiology (pp. 305-323).
Rothenberg, M.
(1981b). The voice source in
singing. In Research Aspects on Singing (Publication #33).
Rothenberg, M.
(1988). Acoustic reinforcement of vocal
fold vibratory behavior in singing. In Vocal Fold Physiology: Voice Production,
Mechanisms and Functions.
Roubeau, B., Chevrie-Muller, C., & Arabia-Guidet, C.
(1987). Electro-glottographic study of the changes of voice
registers. Folia Phoniatrica, 39, 280-289.
Saxon, K.G.,
& Schneider, C.M. (1995). Vocal Exercise Physiology.
Schlümicher-Thier,
J., Zauner, B., Weikert, M.,
Shahidullah, S.,
& Hepper, P.G. (1994). Frequency
discrimination by the fetus. Early Human Development, 36, 13-26.
Shiotani, A.,
Fukuda, H., Kawaida, M., & Kanzaki, J. (1996). Vocal fold vibration in simulated head voice
phonation in excised canine larynges. European Archives of Otolaryngology,
253(6), 356-363.
Shipp, T.
(1975). Vertical laryngeal position
during continuous and discrete vocal frequency change. Journal
of Speech and Hearing Research, 18(4), 707-718.
Shipp, T.,
Lindestad, P.-Å., MacCurtain, F.,
Shipp, T., &
McGlone, R. (1971). Laryngeal dynamics
associated with voice frequency change. Journal of Speech and Hearing Research,
14, 761-768.
Siegel, D.J.
(1999). The Developing Mind: Toward a Neurobiology of Interpersonal Experience.
Smotherman,
W.P., & Robinson, S.R. (1995).
Tracing developmental trajectories into the prenatal period. In T.P. Lecanuet, W.P. Fifer, N.A. Krasnegor,
& W.P. Smotherman (Eds.), Fetal
Development: A Psychobiological Perspective.
Strong, M.S.,
& Vaughan, C.W. (1981). The
morphology of the phonatory organs and their neural control. In K.N. Stevens & M. Hirano (Eds.), Vocal Fold Physiology (pp. 13-22).
Sundberg, J.
(1987). The voice source. In J. Sundberg, The Science of the Singing Voice (pp. 49-92).
Sundberg, J.,
& Kullberg, A. (1998). Voice source
studies of register differences in untrained female voices. Royal
Institute of Technology - Speech,
Music and Hearing Quarterly Progress, (TMH-QPSR), 1-2, 9-17.
Sundberg, J.,
& Högset, C. (1999). Voice source
differences between falsetto and modal registers in countertenors, tenors, and
baritones. Royal Institute of Technology
- Speech, Music and Hearing Quarterly
Progress, (TMH-QPSR), 3-4, 65-74.
Tanaka, K., Kitajima, K., & Kataoka, H. (1997). Effects
of tranglottal pressure change on fundamental frequency of phonation:
Preliminary evaluation of the effect of intraoral pressure change. Folia
Phoniatrica et Logopedica, 49(6), 300-307.
Thurman, L.
(1988). Parental singing during
pregnancy and infancy can assist in cultivating positive bonding and later
development. In Fedor-Freybergh, P.G.,
& Vogel, M.L.V. (Eds.), Prenatal and
Perinatal Psychology and Medicine:
Encounter with the Unborn (pp. 273-282),
Thurman, L.,
Chase, M., & Langness, A.P. (1987).
Reaching the young child through music: Is pre-natal and infant music
education possible? International Music Education, 14, 21-28.
Thurman, L.,
& Welch, G. (2000). Bodymind and Voice: Foundations of Voice
Education (Rev. Ed.).
Timberlake, C.
(1990). Practicae musicae:
Terminological turmoil--the naming of registers. The
National Association of Teachers of Singing Journal, 47(1), 14-26.
Titze,
Titze, I.R.
(1983). The importance of vocal tract
loading in maintaining vocal fold oscillation.
Proceedings from the
Titze, I.R.
(1984). Influences of subglottal
resonance on the primary register transition.
In V.L. Lawrence (Ed.). Transcripts of the Thirteenth Symposium: Care of the Professional Voice
(pp. 130-134).
Titze, I.R.
(1988). A framework for the study of
vocal registers. Journal of Voice, 2(3),183-194.
Titze, I.R.
(1990). Interpretation of the
electroglottographic signal. Journal of Voice, 4(1),1-9.
Titze,
Titze,
Titze,
I.R. (2000c). Generation and propagation
of sound. In I.R. Titze, Principles of Voice Production (pp.
123-148).
Titze, I.R.
(2000d). Vocal registers. In Principles
of Voice Production (pp. 281-310).
Titze, I.R.,
Luschei, E., & Hirano, M. (1989).
Role of the thyroarytenoid muscle in regulation of fundamental
frequency. Journal of Voice, 3, 213-224.
Tom, K., Titze,
I.R., Hoffman, E.A., & Story, B.H. (2001).
Three-dimensional vocal tract imaging and formant structure: Varying
vocal register, pitch, and loudness. Journal of the Acoustical Society of
Van den Berg,
J.W. (1958). Myoelastic-aerodynamic
theory of voice production. Journal of Speech and Hearing Research,
1, 227-244.
van Diense, J.
(1982). On vocal registers. Journal
of Research in Singing, 5(2), 33-39.
Vander, A.J., Sherman, J.H., & Luciano, D.S.
(1994). Human Physiology: The
Mechanisms of Body Functions
(6th Ed.).
Vennard, W.
(1967). Singing: The Mechanism and Technic.
Vennard, W.,
Hirano, M., & Ohala, J. (1970a).
Chest, head and falsetto. The National Association of Teachers of
Singing Bulletin, 27, 30-36.
Vennard, W., Hirano, M., & Ohala, J. (1970b). Laryngeal
synergy in singing. The National Association of Teachers of Singing Bulletin, 27(1),
16-21.
Vilkman, E., Alku, P., & Laukkanen, A.-M.
(1995). Vocal-fold collision mass as a differentiator between
registers in the low-pitch range. Journal of Voice, 9(1), 66-73.
Vrbova, G.,
Gordon, T., & Jones, R. (1994). Nerve-Muscle Interaction.
Webster, D.B.
(1995). Neuroscience of Communication.
Welch, G.F., Sergeant, D.C., & MacCurtain, F.
(1988). Some physical characteristics of the male falsetto
voice. Journal of Voice, 2(2), 151-163.
Welch, G.F.,
Sergeant, D.C., & MacCurtain, F. (1989).
Xeroradiographic-electrolaryngographic analysis of male vocal
registers. Journal of Voice, 3(3), 244-256.
Wilcox, J.
(1935). The Living Voice.
Williams, R.S.,
Garcia-Moll, M., Mellor, J., Salmons, S., & Harlan, W. (1987). Adaptation of skeletal muscle to increased contractile
activity: Expression of nuclear genes encoding mitochondrial proteins. Journal
of Biological Chemistry, 262, 2764-2767.
Wurgler, P.
(1990). A perceptual study of vocal
registers in the singing voices of children. Unpublished Ph.D. dissertation,
The Ohio State University.
Wyke, B.
(1983a). Neuromuscular control systems
in voice production. In D.M. Bless,
& J.H. Abbs (Eds.), Vocal Fold
Physiology: Contemporary Research and Clinical Issues.
Wyke, B.
(1983b). Reflexogenic contributions to
vocal fold control systems. In I.R.
Titze, & R.C. Scherer, (Eds.) (1983).
Vocal Fold Physiology:
Biomechanics, Acoustics and Phonatory Control (pp. 138-141).
Leon Thurman is Specialist Voice Educator
at Fairview Voice Center, Rehabilitation Services, Fairview-University Medical
Center in Minneapolis, Minnesota, USA, [email: [email protected]], and founder and
Development Director of The VoiceCare Network [www.voicecarenetwork.org].
Graham Welch is Professor of Music
Education and Head of the
Axel Theimer is Professor of Voice and
Choral Music at
Carol Klitzke is Speech Pathologist/Voice
Specialist at